Tausta
eukaryoottisissa kromosomeissa on yleisesti satelliittien DNA: n alueita, joissa esiintyy runsaasti tandem-toistoja keskus-ja perisentromeerisessa kromatiinissa. Sentromeerisen heterokromatiinin tiedetään hyvin vakiinnuttavan kinetokorifunktion ja mahdollistavan uskollisen kromosomien erottelun. Perisentrisen satelliitin DNA: n rooli on kuitenkin vähemmän ymmärretty, mikä johtuu erityisesti proteiinien koodauksen ja säilymisen puutteesta eri lajien välillä. Aiemmissa tutkimuksissa on esitetty rooleja vain tietyissä yhteyksissä, mutta ei tiedetä, onko eukaryoottisen satelliitin DNA: lla keskeisempää roolia, varsinkin kun otetaan huomioon sen runsaus genomissa.
Jagannathan, Cummings ja Yamashita seuraavat tuoreessa eLife-tutkimuksessaan kromosenttien muodostumista ja roolia eli useiden kromosomien niputtamista ydinaseiden organisaation ylläpitämiseksi. Ne käsittelevät useita uusia kysymyksiä, kuten miten kaikki Drosophila melanogaster-kromosomit genomissa voidaan niputtaa kromosenttereihin, ja miten useat satelliitit DNAs ja niitä vastaavat DNA: ta sitovat proteiinit voivat moduloida tätä prosessia yhdessä.
Avainlöydökset
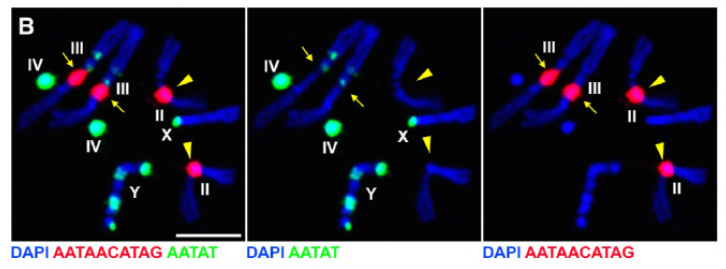
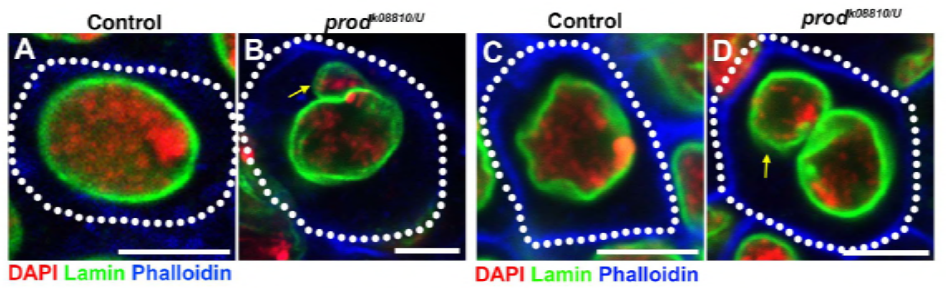
koska heidän edellisessä työssään tunnistettiin proteiini D1, joka oli vastuussa kromosomien X, Y ja 4 yhdistämisestä kromosenttereiksi, ryhmä käänsi huomionsa kromosomeihin 2 ja 3, D. melanogasterin tärkeimpiin autosomeihin, käsitelläkseen sitä, miten kaikki genomin kromosomit voidaan ryhmitellä kromosenttereiksi. Tämä johti siihen, että he havaitsivat autosomeissa pidemmän satelliitin toistuvan, johon Prod (proliferation disrupter) – proteiini sitoutui. Prod: n toiminnan häiritseminen johti mikrotumien muodostumiseen ja lisääntyneeseen DNA-vaurioon, joka johti lopulta solukuolemaan. Tämä nähtiin myös heidän aiemmassa työssään D1-mutanttien kanssa. On kiinnostavaa, että vaikka D1-mutaatiot vaikuttivat itulinjaan, prod-mutaatiot eivät vaikuttaneet, mikä viittaa siihen, että molemmat vaikuttavat eri kudoksiin samankaltaisista kromosentriä muodostavista toiminnoistaan huolimatta. D1 prod-kaksoismutantit eivät kuitenkaan kehity alkiovaiheiden ohi, ja niiden mikrotumat ovat lisääntyneet, minkä vuoksi kromosenttimuodostusta satelliitin DNA: n välityksellä tarvitaan olennaisesti.
Prod: lla on selvästi samanlainen rooli kromosenttereiden muodostamisessa kuin D1: llä. Tekijät ilmentyivät kudoksessa, jossa sitä ei normaalisti ole, ja aiheuttivat kromatiinisäikeiden muodostumisen, jotka muodostavat autosomeja yhdistäviä kromosomialueita. Tämä osoittaa selvästi, että Prod riittää niputtamaan nämä kromosomit yhteen satelliitin DNA: n sitoutumisen kautta, mikä selittää kromosentterien muodostumismekanismin.
tässä vaiheessa Jagannathan ja Cummings et al ovat todenneet Prod-ja D1-klusterikromosomit satelliitin DNA: n avulla. Mutta koska molemmat proteiinit vaikuttavat eri kromosomeihin, niin miten koko genomin kromosomijoukko kapseloituu tumaan? Prod: lla ja D1: llä ei näyttänyt olevan yhteisvaikutuksia immunopresipitaation kautta, mikä viittaa vain heikkoon tai ohimenevään yhteisvaikutukseen. Live-kuvantamisen avulla tekijät löysivät” kiss and run ” – vuorovaikutuksen-D1-ja Prod-foci-kosketukset koskettavat lyhyesti ja sitten erillään, mikä osoittaa kromosenttimuodostuksen dynaamista prosessia. Molemmat proteiinit näyttävät olevan riippuvaisia myös toisen funktionaalisesta läsnäolosta, sillä prod-mutanteilla oli viallinen D1-ryhmitys ytimissä ja päinvastoin. Tämä keskinäisriippuvuus tarjoaa verkoston, jonka avulla genomin kaikki kromosomit voidaan niputtaa yhteen.
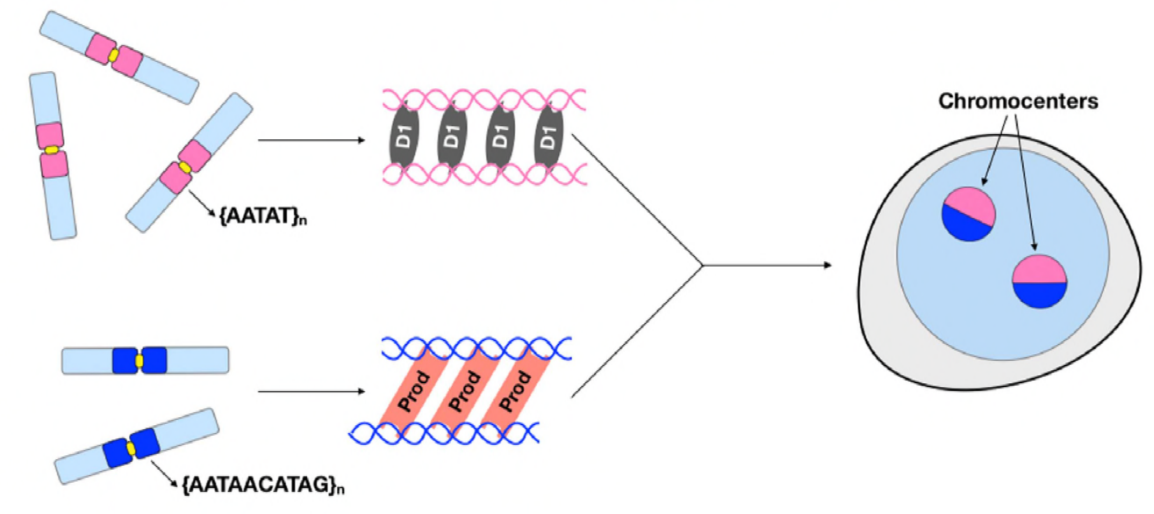
yhteenvetona Jagannathan ja Cummings et al ovat osoittaneet, että Prod ja D1 luovat verkoston, jossa molemmat proteiinit sitovat omat satelliitin DNA-sekvenssinsä, jotta kaikki kromosomit saadaan kromosentraattiin ja lopulta koko genomi pakataan kunnolla tumaan. Heidän tutkimuksensa osoittaa satelliitti-DNA: n merkityksen, ja siinä käsitellään Prod-ja D1-häiriöiden molekyyli-ja solubiologisten seurausten lisäksi myös näiden proteiinien ja satelliitti-DNA: n evolutiivista merkitystä.
kysymykset kirjoittajille
elife – tutkimus osoitti, että D1: n häviäminen saa mikrotumat alkamaan pääytimestä-aiheuttaako prod: n häviäminen mikrotumien muodostumisen samalla tavalla?
ovatko Prod ja D1 vuorovaikutuksessa muiden proteiinien kanssa lukuun ottamatta niiden ohimeneviä ”kiss and run” – vuorovaikutuksia? Tiedetäänkö näiden proteiinien toimivan muissa ydinkestävyyteen ja/tai genomipakkauksiin liittyvissä prosesseissa?
eliöissä, joilla on yli 4 kromosomia, kuinka monta kromosentriä muodostavaa, satelliitin DNA: ta sitovaa proteiinia saattaa olla tarpeen liittää yhteen suurempi genomi, jossa on enemmän kromosomeja?
keskustelussa mainitaan, että D. simulansilla ei ole satelliitin DNA: ta, joka sitoo D. melanogaster Prodin, joten mikä on D. simulansin Prodin kaltainen proteiini ja kuinka samankaltainen se on rakenteeltaan ja toiminnaltaan D. melanogasterin kanssa?
Tags: cell biology, drosophila, fruit flies, genetic, genome packaging, satellite dna
Posted on: 11th December 2018
doi: https://doi.org/10.1242/prelights.6160