fundal
cromozomii Eucarioți au în mod obișnuit regiuni de ADN satelit abundente în repetări tandem la cromatină centromerică și pericentromerică. Heterocromatina centromerică este bine cunoscută pentru a stabili funcția kinetocorului și pentru a permite segregarea fidelă a cromozomilor. Cu toate acestea, rolul ADN-ului satelit pericentric este mai puțin înțeles, în special din cauza lipsei de codificare și conservare a proteinelor între specii. Studiile anterioare au arătat roluri doar pentru contexte specifice, dar nu se știe dacă există un rol mai central pentru ADN-ul satelit eucariot, în special având în vedere abundența sa în genom.
cu această nouă preimprimare, Jagannathan, Cummings și Yamashita urmăresc studiul lor recent eLife care discută despre formarea și rolul cromocenterilor sau gruparea cromozomilor multipli, pentru a menține organizarea nucleară. Ei abordează mai multe întrebări noi, inclusiv modul în care toți cromozomii Drosophila melanogaster dintr-un genom pot fi grupați în cromocenteri și modul în care ADN-urile multiple prin satelit și proteinele lor de legare a ADN-ului corespunzătoare pot modula acest proces împreună.
constatări cheie
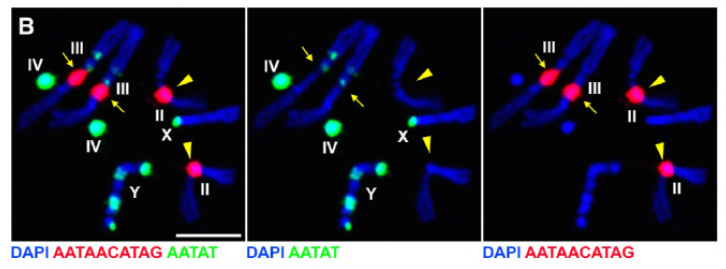
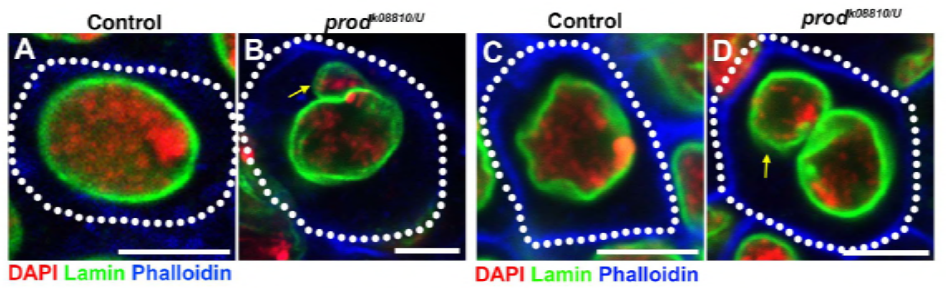
deoarece munca lor anterioară a identificat o proteină, D1, care a fost responsabilă pentru gruparea cromozomilor X, Y și 4 în cromocenteri, grupul și-a îndreptat atenția către cromozomii 2 și 3, autozomii majori din D. melanogaster, pentru a aborda modul în care toți cromozomii din genom pot fi grupați în cromocenteri. Acest lucru i-a determinat să descopere o repetare mai lungă a satelitului pe autozomii legați de proteina Prod (proliferation disrupter). Perturbarea funcției prod a dus la formarea micronucleelor și la creșterea deteriorării ADN-ului, ducând în cele din urmă la moartea celulară. Acest lucru a fost văzut și în lucrarea lor anterioară cu mutanții D1. Interesant este că, în timp ce mutațiile D1 au afectat linia germinală, mutațiile prod nu au făcut-o, sugerând astfel că ambele acționează asupra țesuturilor diferite, în ciuda funcțiilor lor similare de formare a cromocentrului. Cu toate acestea, mutanții dubli D1 prod nu reușesc să se dezvolte dincolo de etapele embrionare și au crescut micronuclei, stabilind cerința esențială a formării cromocentrului prin ADN satelit.
Prod are în mod clar un rol similar cu D1 în formarea cromocentrelor. Autorii au exprimat Prod ectopic într-un țesut în care în mod normal nu este prezent și a provocat formarea de fire de cromatină care stabilesc diferite teritorii cromozomiale care leagă autozomii. Acest lucru arată clar că Prod este suficient pentru a lega acești cromozomi împreună prin legarea ADN-ului său satelit, explicând mecanismul modului în care se formează cromocentrii.
în acest moment, Jagannathan și Cummings și colab au stabilit că cromozomii cluster Prod și D1 prin ADN-ul satelit. Dar din moment ce ambele proteine acționează pe seturi diferite de cromozomi, cum este un întreg set de cromozomi al genomului încapsulat într-un nucleu? Prod și D1 nu par să interacționeze împreună prin imunoprecipitare, sugerând doar o interacțiune slabă sau tranzitorie. Prin imagistica live, autorii au descoperit o interacțiune” kiss and run ” – focarele D1 și focarele Prod ating scurt și apoi se separă, indicând un proces dinamic de formare a cromocentrului. Ambele proteine par să depindă reciproc și de prezența funcțională a celuilalt, deoarece mutanții prod au prezentat clustering defect D1 în nuclee și invers. Această interdependență oferă o rețea pentru a stabili gruparea tuturor cromozomilor dintr-un genom.
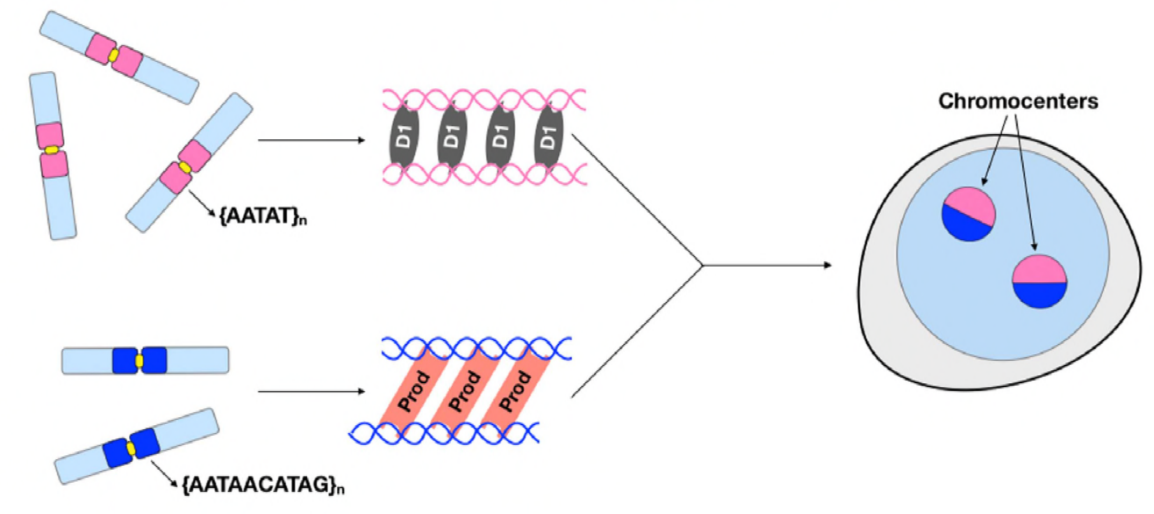
în rezumat, Jagannathan și Cummings și colab au demonstrat că Prod și D1 creează o rețea în care ambele proteine își leagă secvențele de ADN satelit respective pentru a aduce toți cromozomii într-un cromocentru și, în cele din urmă, împachetează întregul genom în mod corespunzător în nucleu. Studiul lor demonstrează importanța ADN-ului satelit, abordând nu numai consecințele biologice moleculare și celulare ale perturbațiilor Prod și D1, ci și semnificația evolutivă a acestor proteine și a ADN-ului satelit.
întrebări pentru autori
studiul elife a arătat că pierderea D1 determină micronuclee să se desprindă de nucleul principal – pierderea prod provoacă formarea micronuclee în același mod?
prod și D1 interacționează cu alte proteine în afară de interacțiunile lor tranzitorii de „sărut și alergare”? Se știe că aceste proteine funcționează în alte procese legate de integritatea nucleară și/sau ambalarea genomului?
în organismele cu mai mult de 4 cromozomi, câte proteine de legare a ADN-ului prin satelit care formează cromocentrul ar putea fi necesare pentru a lega un genom mai mare cu mai mulți cromozomi?
discuția menționează că D. simulans nu are ADN-ul satelit care leagă D. melanogaster Prod, deci care este proteina asemănătoare Prod în D. simulans și cât de asemănătoare este cu D. melanogaster în structură și funcție?
Tags: biologie celulara, drosophila, muste de fructe, genetica, ambalare genomului, ADN-ul prin satelit
postat pe: 11 decembrie 2018
doi: https://doi.org/10.1242/prelights.6160