háttér
az eukarióta kromoszómáknak gyakran vannak műholdas DNS-régiói, amelyek a centromer és a pericentromer kromatin mellett tandem ismétlődésekben bővelkednek. A centromer heterokromatin jól ismert, hogy létrehozza a kinetochore funkciót, és lehetővé teszi a hűséges kromoszóma szegregációt. A pericentrikus műholdas DNS szerepe azonban kevésbé ismert, különösen a fehérjék kódolásának és a fajok megőrzésének hiánya miatt. Korábbi tanulmányok csak meghatározott összefüggésekben mutattak szerepeket, de hogy van-e központi szerepe az eukarióta műholdas DNS-nek, különös tekintettel a genomban való bőségére, nem ismert.
ezzel az új preprinttel Jagannathan, Cummings és Yamashita nyomon követik a legutóbbi Elife tanulmányukat, amely a kromocentrumok kialakulását és szerepét tárgyalja, vagy több kromoszóma összekapcsolását a nukleáris szervezet fenntartása érdekében. Számos új kérdéssel foglalkoznak, beleértve azt is, hogy a genom összes Drosophila melanogaster kromoszómája hogyan köthető össze kromocentrumokban, és hogyan lehet több műholdas DNS és a megfelelő DNS-kötő fehérje együtt modulálni ezt a folyamatot.
főbb megállapítások
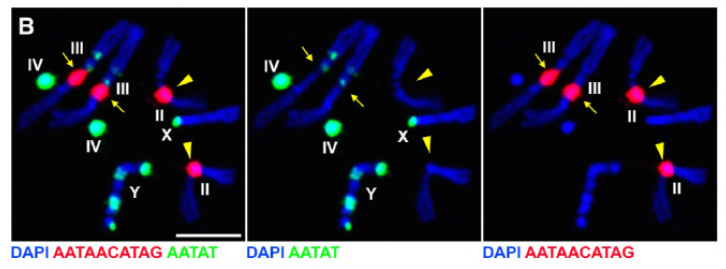
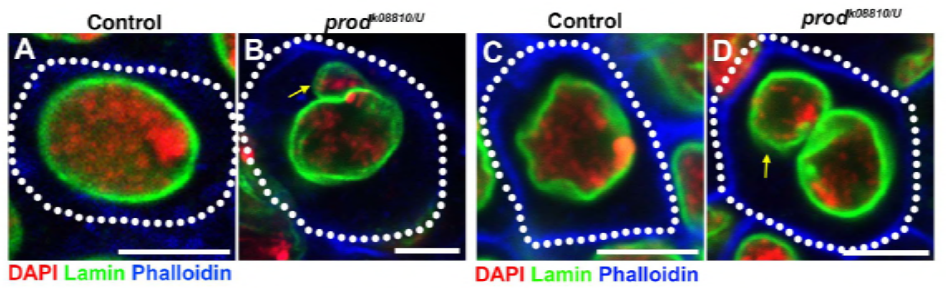
mivel korábbi munkájuk során azonosítottak egy fehérjét, a D1-et, amely felelős volt az X, Y és 4 kromoszómák kromocentrumokba történő kötegeléséért, a csoport a 2-es és 3-as kromoszómákra, a D. melanogaster fő autoszómáira fordította figyelmét, hogy foglalkozzanak azzal, hogy a genom összes kromoszómája hogyan csoportosulhat kromocentrumokba. Ez arra késztette őket, hogy felfedezzenek egy hosszabb műholdas ismétlést az autoszómákon, amelyeket a Prod (proliferation disrupter) fehérje kötött. A prod funkció megzavarása mikronukleuszok képződéséhez és fokozott DNS-károsodáshoz vezetett, ami végül sejthalált eredményezett. Ezt a D1 mutánsokkal végzett korábbi munkájukban is látták. Érdekes módon, míg a D1 mutációk befolyásolták a csíravonalat, a prod mutációk nem, ami arra utal, hogy mindkettő hasonló kromocentrumképző funkcióik ellenére különböző szövetekre hat. A D1 prod kettős mutánsok azonban nem fejlődnek túl az embrionális szakaszokon, és megnövekedtek a mikronukleuszok, ami megalapozza a kromocentrum képződésének alapvető követelményét műholdas DNS-en keresztül.
a Prod egyértelműen hasonló szerepet játszik a D1-hez a kromocentrumok kialakításában. A szerzők ektopikusan fejezték ki a Prod-ot egy olyan szövetben, ahol általában nincs jelen, és kromatin szálak képződését okozták, amelyek különböző kromoszóma területeket hoznak létre, amelyek összekapcsolják az autoszómákat. Ez egyértelműen azt mutatja, hogy a Prod elegendő ahhoz, hogy ezeket a kromoszómákat műholdas DNS-kötésén keresztül összekapcsolja, elmagyarázva a kromocentrumok kialakulásának mechanizmusát.
ezen a ponton Jagannathan és Cummings et al megállapították, hogy a Prod és a D1 klaszter kromoszómákat műholdas DNS-en keresztül. De mivel mindkét fehérje különböző kromoszómakészletekre hat, hogyan van egy teljes genom kromoszómakészlete kapszulázva egy magba? Úgy tűnt, hogy a Prod és a D1 nem lép kölcsönhatásba immunprecipitáció útján, ami csak gyenge vagy átmeneti kölcsönhatásra utal. Az élő képalkotás során a szerzők felfedezték a” kiss and run ” interakciót – a D1 gócok és a Prod gócok röviden érintkeznek, majd elválnak, jelezve a kromocenter kialakulásának dinamikus folyamatát. Úgy tűnik, hogy mindkét fehérje kölcsönösen függ a másik funkcionális jelenlététől is, mivel a prod mutánsok hibás D1 csoportosulást mutattak az atommagokban és fordítva. Ez az egymásrautaltság hálózatot biztosít a genomban lévő összes kromoszóma összekapcsolásának megállapításához.
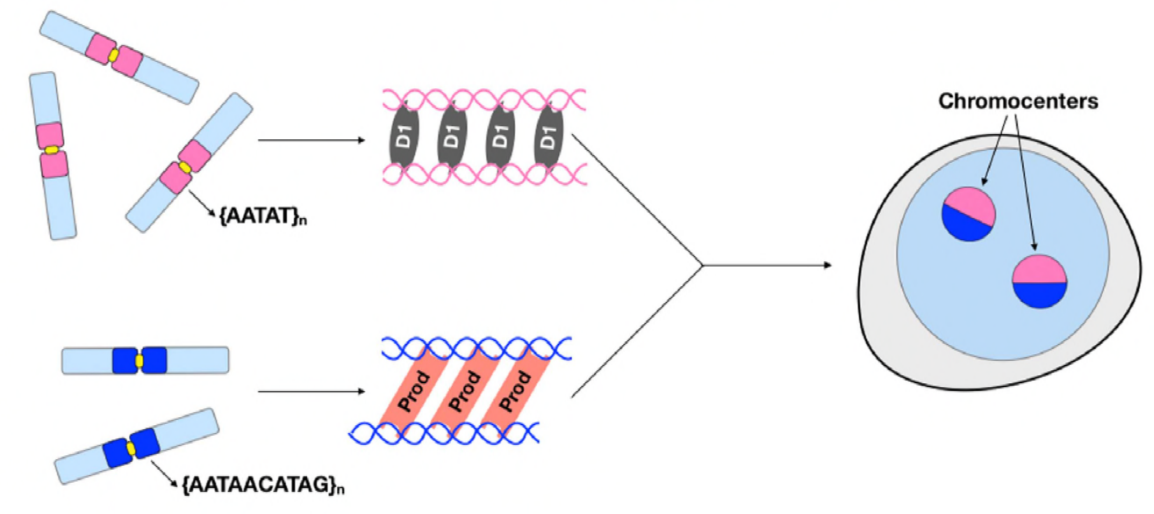
összefoglalva, Jagannathan és Cummings et al kimutatták, hogy a Prod és a D1 olyan hálózatot hoz létre, amelyben mindkét fehérje megköti a saját műholdas DNS-szekvenciáját annak érdekében, hogy az összes kromoszómát egy kromocentrumba hozza, és végül a teljes genomot megfelelően csomagolja a magba. Tanulmányuk bizonyítja a műholdas DNS fontosságát, nemcsak a Prod és a D1 perturbációk molekuláris és Sejtbiológiai következményeivel foglalkozik, hanem ezen fehérjék és a műholdas DNS evolúciós jelentőségével is.
kérdések a szerzőkhöz
az elife tanulmány kimutatta, hogy a D1 veszteség a mikronukleuszok rügyét okozza a fő magból – a prod veszteség ugyanúgy okozza a mikronukleuszok képződését?
a Prod és a D1 kölcsönhatásba lépnek-e más fehérjékkel, kivéve az átmeneti “kiss and run” kölcsönhatásokat? Ismertek-e ezek a fehérjék a nukleáris integritással és/vagy a genomcsomagolással kapcsolatos egyéb folyamatokban?
a 4-nél több kromoszómával rendelkező organizmusokban hány kromocentrikus, műholdas DNS-kötő fehérje szükséges ahhoz, hogy egy nagyobb genomot több kromoszómával kössön össze?
a vita megemlíti, hogy a D. simulans nem rendelkezik a D. melanogaster Prod-hoz kötődő műholdas DNS-sel, tehát mi a D. simulans Prod-szerű fehérje, és mennyire hasonlít a D. melanogaster szerkezetére és funkciójára?
címkék: sejtbiológia, drosophila, gyümölcs legyek, genetika, Genom csomagolás, műholdas DNS
feladta: 11th December 2018
doi: https://doi.org/10.1242/prelights.6160