Bakgrunn
Eukaryote kromosomer har ofte regioner av satellitt-DNA rikelig i tandem gjentar ved centromeric og pericentromeric kromatin. Centromeric heterochromatin er godt kjent for å etablere kinetochore funksjon og tillate trofast kromosom segregering. Imidlertid er rollen til perikentrisk satellitt-DNA mindre forstått, særlig på grunn av mangel på proteinkoding og bevaring på tvers av arter. Tidligere studier har vist roller for spesifikke sammenhenger bare, men om det er en mer sentral rolle for eukaryotisk satellitt-DNA, spesielt gitt sin overflod i genomet, er ikke kjent.
Med Denne nye preprint, Jagannathan, Cummings, Og Yamashita følge opp på deres siste eLife studie diskuterer dannelsen og rolle chromocenters, eller bunting av flere kromosomer, for å opprettholde kjernefysisk organisasjon. De adresserer flere nye spørsmål, inkludert hvordan Alle Drosophila melanogaster kromosomer i et genom kan buntes i kromocentre, og hvordan flere satellitt-Dnaer og deres tilsvarende DNA-bindende proteiner kan modulere denne prosessen sammen.
Nøkkelfunn
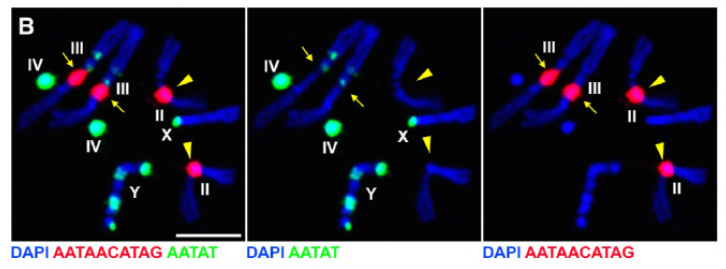
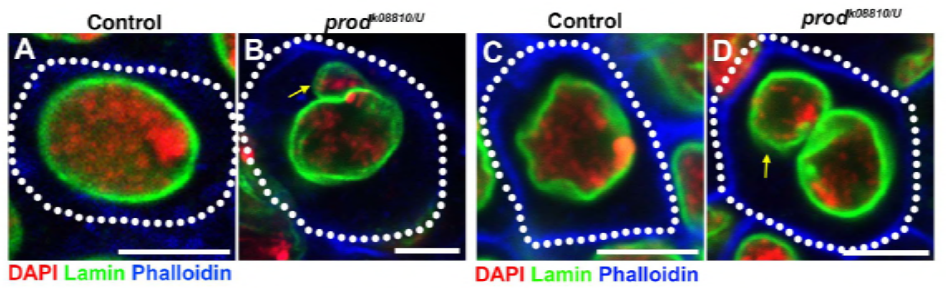
Siden deres tidligere arbeid identifiserte et protein, D1, som var ansvarlig for bunting av kromosomer X, Y og 4 i kromocentre, viste gruppen oppmerksomheten mot kromosomer 2 og 3, de viktigste autosomene I d. melanogaster, for å ta opp hvordan alle kromosomer i genomet kan grupperes i kromocentre. Dette førte dem til å oppdage en lengre satellitt gjenta på autosomer som var bundet Av Prod (proliferation disrupter) protein. Forstyrrende prod-funksjon førte til mikronukleidannelse og økt DNA-skade, noe som til slutt resulterte i celledød. Dette ble også sett i deres tidligere arbeid Med d1 mutanter. Interessant, Mens D1-mutasjoner påvirket kimlinjen, gjorde prod-mutasjoner ikke, noe som tyder på at begge virker på forskjellige vev til tross for deres lignende kromocenterdannende funksjoner. D1 prod doble mutanter ikke klarer å utvikle seg forbi embryonale stadier og har økt mikronuclei, etablere det grunnleggende kravet om chromocenter dannelse via satellitt-DNA.
Prod har tydeligvis en lignende rolle Som D1 i dannelsen av kromocentre. Forfatterne uttrykte prod ektopisk i et vev hvor det normalt ikke er tilstede og forårsaket dannelsen av kromatintråder som etablerer forskjellige kromosomområder som forbinder autosomene. Dette viser tydelig At Prod er tilstrekkelig til å pakke disse kromosomene sammen gjennom sin satellitt-DNA-binding, og forklarer mekanismen for hvordan kromocentre dannes.
På dette tidspunktet Har Jagannathan og Cummings et al etablert At Prod Og D1 klynger kromosomer gjennom satellitt-DNA. Men siden begge proteiner virker på forskjellige sett med kromosomer, hvordan er et helt genom sett med kromosomer innkapslet i en kjerne? Prod og D1 så ikke ut til å interagere sammen gjennom immunopresipitasjon, noe som bare tyder på en svak eller forbigående interaksjon. Gjennom levende bildebehandling oppdaget forfatterne en» kiss and run » – interaksjon-D1 foci og Prod foci touch kort og deretter skille, noe som indikerer en dynamisk prosess med kromocenterdannelse. Begge proteinene ser ut til å være gjensidig avhengig av den andres funksjonelle tilstedeværelse også, siden prod mutanter viste defekt D1 clustering i kjerner og omvendt. Denne gjensidige avhengigheten gir et nettverk for å etablere bunting av alle kromosomer i et genom.
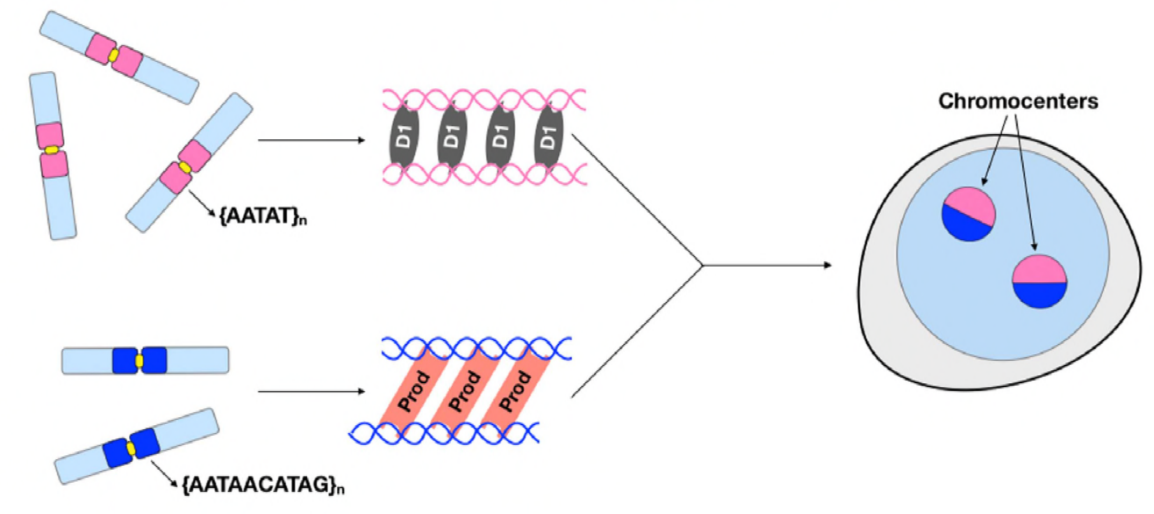
I sammendraget Har Jagannathan og Cummings et al vist At Prod og D1 skaper et nettverk hvor Begge proteiner binder sine respektive satellitt-DNA-sekvenser for å bringe alle kromosomer inn i et kromocenter, og til slutt pakke hele genomet riktig inn i kjernen. Deres studie demonstrerer betydningen av satellitt-DNA, og adresserer ikke bare de molekylære og cellebiologiske konsekvensene Av prod-Og D1-forstyrrelser, men også den evolusjonære betydningen av disse proteinene og satellitt-DNA også.
spørsmål til forfatterne
elife-studien viste at D1-tap fører til at mikronuklei knopper av fra hovedkjernen-forårsaker prod-tap mikronukleidannelse på samme måte –
interagerer Prod Og D1 med andre proteiner bortsett fra deres forbigående «kyss og løp» – interaksjoner? Er disse proteinene kjent for å fungere i andre prosesser relatert til kjerneintegritet og / eller genomemballasje?
i organismer med mer enn 4 kromosomer, hvor mange kromocenterdannende, SATELLITT-DNA-bindende proteiner kan være nødvendige for å bunte et større genom med flere kromosomer?
diskusjonen nevner At D. simulans ikke har satellitt-DNA som binder d. melanogaster Prod, så hva Er Det Prod-lignende proteinet I D. simulans og hvordan ligner Det På d. melanogaster i struktur og funksjon?
Stikkord: cellebiologi, drosophila, fruktfluer, genetikk, genom emballasje, satellitt dna
Postet på: 11 desember 2018
doi: https://doi.org/10.1242/prelights.6160