tarttuva geneettinen materiaali varastoituu solujen sisälle kromosomeiksi kutsuttuihin rakenteisiin, joiden molemmissa päissä on toistuva sekvenssi, joka tunnetaan telomeerina. Erikoistuneet proteiinit sitoutuvat näihin sekvensseihin muodostaen suojaavan ”korkin”, joka suojaa kromosomia ja estää sitä fuusioitumasta muiden kromosomien kanssa. Telomeraasientsyymi auttaa myös kromosomien ylläpitämisessä lisäämällä telomeerien päihin toistuvia DNA-jaksoja.
yksi laajimmin tutkituista rajausmolekyyleistä on proteiini nimeltä Cdc13, joka sitoutuu tietyntyyppiseen yksijuosteiseen DNA: han orastavassa hiivassa ja muodostaa kompleksin kahden muun proteiinin (Stn1 ja Ten1) kanssa, jotka värittävät telomeraasia (Wellinger ja Zakian, 2012). Useat todisteet viittaavat siihen, että tämä CST-kompleksi rekrytoi myös DNA: n replikaatioentsyymin nimeltä primaasi-Pola ja voi säädellä tämän entsyymin toimintaa kromosomien päissä sekä muissa genomin paikoissa (Giraud-Panis et al., 2010; Price et al., 2010; Barbero Barcenilla and Shippen, 2019).
samanlaisia komplekseja on havaittu myös muissa eukaryooteissa, kuten nisäkkäissä, jotka sisältävät STN1: tä, Ten1: tä ja toista ctc1-nimistä proteiinia CST-kompleksissaan (Giraud-Panis et al., 2010; Price et al., 2010). CST-kompleksin eri proteiinien roolien selvittäminen on kuitenkin osoittautunut haastavaksi, koska solut, joilta puuttuu vain yksi näistä proteiineista, kamppailevat selviytyäkseen (Kuva 1a). Nyt Elife, Jin-Qiu Zhou ja työtovereiden Kiinan tiedeakatemian ja ShanghaiTech University-mukaan lukien Zhi-Jing Wu ensimmäisenä kirjoittajana-raportoivat tulokset kokeista, jotka auttavat parantamaan ymmärrystämme CST monimutkainen (Wu et al., 2020).
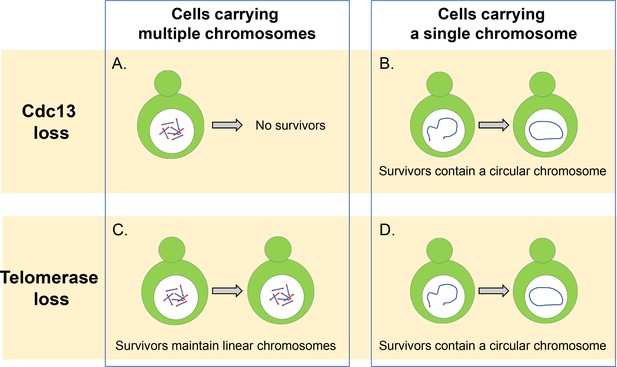
Cdc13 ja telomeraasi ovat välttämättömiä lineaaristen kromosomien säilyttämiseksi.
(a) hiivasolut, joilla on useita lineaarisia kromosomeja, tarvitsevat cdc13-korkkiproteiinia suojellakseen telomeerejaan ja estääkseen kromosomien sulautumisen. Ilman tätä proteiinia nämä solut eivät voi selviytyä. B) solut, joilla on yksi lineaarinen kromosomi, voivat selviytyä ilman Cdc13: a liittämällä kromosominsa päät yhteen muodostaen pyöreän renkaan. C) ilman telomeraasientsyymiä solut, joilla on useita lineaarisia kromosomeja, pystyvät selviytymään käyttämällä DNA: n rekombinaatioreittejä, jotka voivat monistaa telomeerijaksoa tai kromatiini-ja telomeerijakson välissä olevia DNA-segmenttejä. D) solut, joilla on yksi lineaarinen kromosomi, selviävät telomeraasin häviämisestä fuusioitumalla ympyränmuotoiseksi kromosomiksi homologisella rekombinaatiolla, kuten tapahtuu soluissa, joilta puuttuu proteiini Cdc13.
Kuvasaldo: Constance Nugent ja Katsunori Sugimoto.
ensin Wu ym. tutkittiin, miten CST-kompleksin poistaminen vaikutti orastavan hiivakannan elinkelpoisuuteen, jossa kaikki sen 16 kromosomia fuusioitiin yhteen muodostaen yhden ympyränmuotoisen kromosomin (Shao et al., 2019). He havaitsivat, että CST: n poistaminen ei estänyt soluja lisääntymästä eikä johtanut useampiin solukuolemiin, vaikka ympyränmuotoinen kromosomi sisälsi toistuvia telomeerijaksoja. Näyttää siis siltä, että CST-kompleksin tärkein tehtävä on ylläpitää lineaarisia kromosomeja ja estää kromosomeja fuusioitumasta muiden kromosomien kanssa, eikä se ole välttämätön sisäisten telomeerisekvenssien replikoinnille.
pyöreän renkaan muodostamisen lisäksi orastavan hiivan 16 kromosomia voidaan myös sulauttaa yhteen muodostaen yhden lineaarisen kromosomin (Shao et al., 2018). Wu ym. havaittiin, että CST-kompleksin poistaminen vähensi huomattavasti näiden solujen elinkelpoisuutta, mutta jotkut näistä soluista pystyivät selviytymään fuusioimalla niiden lineaarisen kromosomin päät muodostaen pyöreän renkaan (Kuva 1b). CST-kompleksin eri proteiineja koodaavien geenien yksittäin poistaminen paljasti, että cdc13: sta puuttuvat solut fuusioituvat nopeammin kuin stn1: n ja Ten1: n geenit puuttuvat. Tämä viittaa siihen, että Cdc13: lla on hallitseva rooli kromosomien fuusion estämisessä ja että Stn1 ja Ten1 edistävät telomeerien suojaamista Cdc13: sta riippumatta. Mekanismin yksityiskohdat ovat kuitenkin vielä epäselviä ja vaativat lisätutkimuksia.
villin tyypin soluissa, joissa on useita kromosomeja, on harvinaista löytää fuusioituneita tai pyöreitä kromosomeja, vaikka telomeraasin aktiivisuus olisi vaarantunut: tämä johtuu siitä, että solut voivat laajentaa ja ylläpitää telomeerejä käyttämällä homologisesti ohjattua rekombinaatiota, joka korjaa kaksijuosteisia katkoksia DNA: ssa (Kuva 1c). Kuitenkin, Wu et al. havaittiin, että kromosomien määrän vähentäminen johti siihen, että soluissa havaittiin enemmän fuusioita, joista puuttui telomeraasientsyymi. Tämä viittaa siihen, että kromosomien määrän vähentäminen lisää todennäköisyyttä, että solut pystyvät tuottamaan ympyröityjä kromosomeja ja selviämään telomeraasin häviämisestä.
ajateltiin, että yksiköllisen lineaarisen kromosomin kahden pään sulauttaminen riippuisi DNA: n korjausreitistä, jota kutsutaan nonhomologiseksi End-Jointing (Nhej) – reitiksi (Haber, 2016). Kuitenkin, Wu et al. on osoitettu, että ilman telomeraasia kromosomien fuusio riippui Rad52: sta, jolla on ratkaiseva rooli orastavan hiivan DNA-katkosten homologisessa rekombinaatiossa (Kuva 1D). On mahdollista, että tässä tutkimuksessa käytetyt solut tukeutuvat rad52-kulkureittiin kromosomien kiertokulussa, koska yksittäisessä kromosomissa on käänteinen telomeerijakso lähellä kromosomin toista päätä. Jos tällainen sekvenssi poistetaan, solut saattavat läpikäydä päästä päähän fuusion ihmisen soluissa yleisemmän nhej-reitin kautta (Palm and de Lange, 2008). Lisäkokeet osoittivat, että tämä havainto ei johtunut nhej: n aktiivisuuden vähenemisestä ja että tämä reitti pystyy sulattamaan lineaarisia plasmideja orastavissa hiivasoluissa.
Wun ym. tarjoaa uusia oivalluksia siitä, miten kromosomit sulautuvat yhteen ja miten telomeerit säilyvät itsenäisesti telomeraasientsyymistä. Lisäksi tämän tutkimuksen havainnot saattavat ylittää hiivan ja parantaa ymmärrystämme erilaisista ihmisen lääketieteellisistä oireyhtymistä, jotka johtuvat kromosomien päiden fuusioitumisesta rengasmuotojen muodostamiseksi (Pristyazhnyuk and Menzorov, 2018).