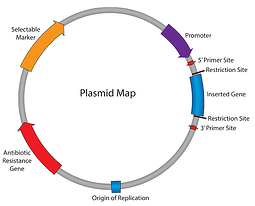
 tähän mennessä meidän plasmidit 101 sarja olemme työskennelleet läpi plasmidikartta: antibioottiresistenssi, alkuperä replikaation, ja niin edelleen. Tähän asti voimme kopioida plasmidimme ja varmistaa, että solut säilyttävät sen; seuraava askel on saada plasmidi ilmaisemaan kiinnostavan geenimme. Syötä promoottori. elementti, joka aloittaa transkription RNA: han.
tähän mennessä meidän plasmidit 101 sarja olemme työskennelleet läpi plasmidikartta: antibioottiresistenssi, alkuperä replikaation, ja niin edelleen. Tähän asti voimme kopioida plasmidimme ja varmistaa, että solut säilyttävät sen; seuraava askel on saada plasmidi ilmaisemaan kiinnostavan geenimme. Syötä promoottori. elementti, joka aloittaa transkription RNA: han.
käytännössä termi ”promoottori” kuvaa promoottorin (RNA-polymeraasisitoutumiskohdan) ja operaattoreiden (vastealkioiden) yhdistelmää. Promoottorit ovat noin 100-1000 emäsparin pituisia ja niitä tavataan kohdegeeninsä yläjuoksulla. Promoottorin alueen sekvenssi ohjaa RNA-polymeraasin ja transkriptiotekijöiden sitoutumista, joten promoottoreilla on suuri rooli sen määrittämisessä, missä ja milloin kiinnostavan geenisi ilmaistaan.
RNA-polymeraasi(t)
RNA transkriboidaan DNA: sta RNA-polymeraasin (rnap) avulla. Bakteereissa tämä tapahtuu yhden entsyymin avulla; eukaryooteilla on kuitenkin muliple polymeraaseja, jotka kukin vastaavat tietystä RNAs-osajoukosta. Tämän erityisyyden saavuttamiseksi eukaryoottinen RNAP voi tunnistaa ja sitoutua tiettyihin promoottorielementteihin. Tämä tarkoittaa sitä, että plasmidin selkärangassa olevan promoottorin on oltava yhteensopiva sen RNA-tyypin kanssa, joka on tehtävä: jos haluat mRNA: n (geeniekspressioon), sinun on käytettävä rnap II-promoottoria, kun taas pienet RNA: t (kuten shRNA) transkriboidaan RNAP III-promoottoreista. Tämä viesti sisältää promoottorit yleiseen RNAP II ja RNAP III transkriptio; kuitenkin käyttämällä virus LTRs kuin RNAP II promoottorit on yleisesti käytetty lentiviral ja retroviral konstruktioita ja keskustelemme näistä tulevaisuudessa postitse virusvektorin osia.
Promoottorispesifisyys
sen lisäksi, että valitset promoottorin RNA-transkription perusteella, sinun on myös varmistettava, että plasmidissasi on promoottori, joka soveltuu toimimaan isäntäorganismissasi. Koska transkriptiokoneisto eroaa solutyypeistä tai eliöistä, promoottoreiden on oltava samalla tavalla vaihtelevia. Bakteerien promoottorit toimivat vain prokaryoottisoluissa ja tyypillisesti vain samassa tai läheistä sukua olevassa lajissa, josta ne ovat peräisin. Samoin erilaiset eukaryoottisolutyypit (nisäkäs, hiiva, kasvit jne.) vaativat ainutlaatuisia promoottoreita, ja crossovereita on hyvin vähän. Yleisesti ottaen bakteerien promoottorit ovat vähemmän monimuotoisia ja monimutkaisia, ja niissä on vähemmän osia kuin eukaryoottisissa soluissa. Jotkut promoottorit ovat perustuslaillisesti aktiivisia ja jatkuvasti, kun taas toisia valvotaan tarkemmin. Säännellyt promoottorit saattavat toimia vain tietyissä kudoksissa tai tiettyinä kehitysaikoina, tai ne voidaan kytkeä päälle tai pois päältä halutulla kemikaalilla, lämmöllä tai valolla. Solussa promoottoreita ohjaavat vielä muut säätelytekijät: tehostajat, rajaelementit, eristeet ja äänenvaimentimet; transkription ”vuotavuutta” voi kuitenkin esiintyä. Tämä ei yleensä ole suuri ongelma soluille, mutta se voi sekoittaa tutkimustuloksia tai jopa tappaa soluja, jos kiinnostava geeni on myrkyllinen. Tämän torjumiseksi tutkijat ovat luoneet synteettisiä promoottoreita, jotka sisältävät tyypillisesti jonkin yhdistelmän muita promoottoreita ja ovat yleensä tiukemmin säänneltyjä.
eukaryoottien ja prokaryoottien Yleiset promoottorit
alla on kaksi viitetaulukkoa, joissa luetellaan joitakin yleisimpiä bakteerien ja nisäkkäiden promoottoreita. Nämä listat eivät suinkaan ole tyhjentäviä, mutta niiden pitäisi olla hyvä paikka aloittaa, kun yrität valita täydellisen promoottorin!
eukaryoottiset promoottorit
| promoottori | ensisijaisesti käytetty | RNA-transkriptio | kuvaus | lauseke | muita näkökohtia |
| CMV | yleinen lauseke | mRNA | ihmisen sytomegaloviruksen vahva nisäkäsekspression promoottori | konstitutiivinen | saattaa sisältää tehostumisalueen. Voidaan vaientaa joissakin solutyypeissä. |
| EF1a | yleinen ilmentymä | mRNA | vahva nisäkkäiden ilmentymä ihmisen venymätekijästä 1 alfa | konstitutiivinen | pyrkii antamaan johdonmukaisen ilmentymän solutyypistä tai fysiologiasta riippumatta. |
| SV40 | yleinen lauseke | mRNA | nisäkäsekspression promoottori simian vakuolating virus 40 | konstitutiivinen | voi sisältää tehostajan. |
| PGK1 (ihminen tai hiiri) | yleinen ilmentymä | mRNA | Nisäkäspromoottori fosfoglyseraattikinaasigeenistä. | konstitutiivinen | laajalle levinnyt ilmentymä, mutta voi vaihdella solutyypeittäin. Pyrkii vastustamaan promoottori alas säätelyä metylaation tai deasetylaation vuoksi. |
| Ubc | yleinen lauseke | mRNA | Nisäkäspromoottori ihmisen ubikitiini-C-geenistä | konstitutiivinen | kuten nimestä voi päätellä, tämä promoottori esiintyy kaikkialla. |
| ihmisen beeta-aktiini | yleinen ilmentymä | mRNA | nisäkäspromoottori beeta-aktiinigeenistä | konstitutiivinen | Ubikvitatiivinen. Kanaversiota käytetään yleisesti promoottorihybrideissä. |
| CAG | yleinen lauseke | mRNA | vahva hybridinisäkkäiden promoottori | konstitutiivinen | Sisältää CMV: n tehostajan, kanan beeta-aktiinin promoottorin ja kanin beeta-globiinin splice-hyväksymistyypin. |
| TRE | yleinen lauseke | mRNA | Tetrasykliinivasteelementin promoottori | indusoituva Tetrasyliinin tai sen johdannaisten kanssa. | sisältää tyypillisesti minimaalisen promoottorin, jolla on alhainen perusaktiivisuus ja useita tetrasykliinioperaattoreita. Transkriptio voidaan kytkeä päälle tai pois päältä riippuen siitä, mitä tet-transaktivaattoria käytetään. |
| UAS | yleinen lauseke | mRNA | Drosophila promoottori, joka ylläpitää Gal4: n sitoutumiskohtaa | spesifinen | edellyttää gal4-geenin läsnäoloa promoottorin aktivoimiseksi. |
| Ac5 | yleinen ilmaisu | mRNA | vahva hyönteisten promoottori Drosophila aktiinin 5c-geenistä | konstitutiivinen | käytetään yleisesti Drosophilan ilmentymisjärjestelmissä. |
| Polyhedriini | yleinen lauseke | mRNA | vahva baculovirus | konstitutiivinen | käytetään yleisesti hyönteissolujen ilmaisujärjestelmissä. |
| CaMKIIa | Optogenetiikan geeniekspressio | mRNA | Ca2+/kalmoduliiniriippuvainen proteiinikinaasi II-promoottori | spesifinen | hermosolu – /CNS-ekspressioon käytetty. Kalsiumin ja kalmoduliinin moduloima. |
| GAL1, 10 | yleinen ilmaisu | mRNA | hiivan viereiset, toisistaan poikkeavasti litteroidut promoottorit | indusoituvat galaktoosin kanssa; repressoituvat glukoosin kanssa | voidaan käyttää itsenäisesti tai yhdessä. Säännellään GAL4 ja GAL 80. |
| TEF1 | yleinen lauseke | mRNA | hiivan transkription venymätekijän promoottori | konstitutiivinen | analoginen nisäkkäiden EF1a-promoottorin kanssa. |
| GDS | yleisilme | mRNA | vahva hiiva-ekspression promoottori glyseraldehydin 3-fosfaattidehydrogenaasista | konstitutiivinen | erittäin vahva, jota kutsutaan myös tdh3: ksi tai GAPDH: ksi. |
| ADH1 | yleisilme | mRNA | alkoholidehydrogenaasin Hiivapromoottori i | etanolin tukahduttama | täyspitkä versio on voimakas ja voimakas. Typistetyt promoottorit ovat konstitutiivisia, joilla on alempi ilmaisu. |
| CaMV35S | yleinen ilmaisu | mRNA | vahva kukkakaalin mosaiikkiviruksen aiheuttama kasvinedistäjä | konstitutiivinen | Aktiivinen dikoteissa, vähemmän aktiivinen monokokeissa, jonkin verran aktiivinen eläinsoluissa. |
| Ubi | yleinen ilmentymä | mRNA | maissin ubikitiinigeenin kasvien promoottori | konstitutiivinen | antaa kasveissa suuren ilmentymän. |
| H1 | pieni RNA-ekspressio | shRNA | ihmisen polymeraasi III RNA-promoottorin | konstitutiivinen | ilmentymä voi olla hieman pienempi kuin U6. Se voi ilmentyä paremmin hermosoluissa. |
| U6 | pieni RNA-ekspressio | shRNA | ihmisen U6: sta käytetään myös pientä ydinpromoottoria | Konstitutiivista | hiiren U6: ta, mutta se saattaa olla tehottomampi. |
prokaryoottiset promoottorit
| promoottori | ensisijaisesti käytetty | kuvaus | ilmaisu | muita näkökohtia |
| T7 | in vitro transkriptio/ yleinen lauseke | promoottori T7-bakteriofagista | konstitutiivinen, mutta vaatii T7-RNA-polymeraasin. | in vitro-transkriptioon käytettynä promoottori ajaa joko sense – tai antisense-transkriptiota riippuen sen suuntautumisesta geeniisi. |
| T7lac | korkeat geeniekspressioarvot | promoottori T7 bakteriofagista ja lac-operaattorit | mitätön tyviekspressio, kun sitä ei indusoida. Vaatii T7 – RNA-polymeraasin, jota ohjaa myös lac-operaattori. Voidaan indusoida IPTG: llä. | esiintyy yleisesti pET-vektoreilla. Lac-toimijat sääntelevät sitä hyvin tiukasti. Hyvä moduloimaan geeniekspressiota vaihtelevien induktoripitoisuuksien avulla. |
| Sp6 | in vitro transkriptio/ yleinen lauseke | promoottori SP6 bakteriofagista | konstitutiivinen, mutta vaatii SP6 RNA polymeraasin. | SP6-polymeraasilla on suuri processivity. Kun käytetään in vitro transkriptio, promoottori ajaa joko sense tai antisense transkriptio riippuen sen suuntautuminen geeni. |
| araBAD | yleinen ilmaisu | arabinoosin metabolisen operonin promoottori | Indusoitavissa arabinoosin ja tukahdutetun kataboliitin tukahduttamisella glukoosin läsnä ollessa tai anti-indusoijan fukoosin kilpailullisella sitomisella | heikompi. Esiintyy yleisesti pBAD-vektoreissa. Hyvä nopeaan säätelyyn ja matalaan tyviekspressioon; ei kuitenkaan sovellu hyvin geeniekspression modulointiin vaihtelevien induktoripitoisuuksien avulla. |
| trp | Korkea geenin ekspressio | promoottori E. coli-tryptofaani operoni | Repressiivinen | sammuu, kun solujen tryptofaanipitoisuus on suuri. |
| lac | yleinen lauseke | promoottori lac operonista | konstitutiivinen lac repressorin (lacI tai lacIq) poissaolossa. Voidaan indusoida IPTG: llä tai laktoosilla. | vuotava promoottori, jolla on hieman heikko ilmaisu. lacIq-mutaatio lisää repressorin 10x ilmentymistä, mikä kiristää Lac-promoottorin säätelyä. Hyvä moduloimaan geeniekspressiota vaihtelevien induktoripitoisuuksien avulla. |
| Ptac | yleinen ilmaisu | Lac: n ja trp: n Hybridipromoottori | säännelty kuten lac: n promoottori | Sisältää -35 aluetta trpB: stä ja -10 aluetta lac: stä. Erittäin tiukka sääntely. Hyvä moduloimaan geeniekspressiota vaihtelevien induktoripitoisuuksien avulla. Yleensä parempi ilme kuin lac yksin. |
| pL | Korkea geenin ekspressio | bakteriofagi lambda | promoottori voi olla lämpötilan säätelevä | usein yhdessä lämpötilaherkän cI857-repressorin kanssa. |
| T3 | in vitro transkriptio / yleinen lauseke | promoottori T3-bakteriofagista | konstitutiivinen, mutta vaatii T3-RNA-polymeraasin | in vitro-transkriptioon käytettynä promoottori ajaa joko sense-tai antisense-transkriptiota riippuen sen suuntautumisesta geeniin |
vaikka tämä lista on hyvä paikka aloittaa, yllä olevissa taulukoissa ei pureuduta tutkijoiden käytettävissä oleviin kudoksiin tai kehityskohtaisiin edistäjiin. Plasmideja käytetään usein terapeuttiseen käyttöön, ja näissä tapauksissa on tärkeää tunnistaa oikeat kudoskohtaiset promoottorit, kuten NIH: n tutkijat ovat kuvanneet täällä.
Huom: A. Max Juchheim osallistui tämän artikkelin kirjoittamiseen.