merkitys
kiinnostus circrnoja kohtaan kasvaa, koska ne vaikuttavat biomarkkerin potentiaalin lisäksi moniin biologisiin prosesseihin ja sairauksiin. Ne havaitaan pääasiassa lukemalla, jotka kartoittavat niiden backsplicing junctionia. Tästä huolimatta circrnat eivät ole enää ainoita transkriptejä, jotka sisältävät tällaisen liitoksen, koska viimeaikaiset tutkimukset ovat osoittaneet, että ympyränmuotoiset Dnat ovat yleisiä ja voidaan transkriptoida, jolloin transkriptit jäljittelisivät circRNA-signaalia. Siksi tämä uudentyyppinen kimeerinen transkriptio voi muuttaa tapaa, jolla circRNA-analyysi tehdään, ja vaikuttaa joihinkin jo raportoituihin tuloksiin.
ovatko ympyränmuotoiset Rnat ainoat kimeeriset transkriptit?
ympyränmuotoinen RNA (circRNAs) löydettiin uudelleen muutama vuosi sitten ei-kanonisesti pyöräytettyinä RNA-muotoina, joita esiintyy eri eliöissä, myös ihmisissä (Salzman et al., 2012; Jeck ym., 2013). Ne ovat kovalenttisesti suljettuja transkripteja, jotka on muodostettu RNA: n takaisinkytkentätapahtuman kautta, jossa alavirtaan tulevan eksonin splice-luovuttaja liittyy ylävirtaan splice-vastaanottimeen, mikä johtaa kovalenttisesti suljettuihin transkripteihin, joille on ominaista back-splicing-liitoksen esiintyminen, joka tekee circrnat erotettaviksi lineaarisista vastineistaan (Kuva 1a) (Zhang et al., 2016; Wilusz, 2018).
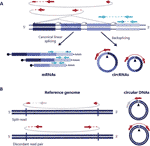
Kuva 1 sekä ympyränmuotoisen RNA: n että DNAs: n havaitseminen perustuu liitoskohtaan ulottuviin lukuihin. (A) Backspliced liittyminen eksonien ja kanoninen eksoni liittyminen on kuvattu ei-jatkuva ja jatkuva linjat vastaavasti aiheuttaa circRNAs ja mRNAs. Pari-end lukee ulottuu backspliced liittymissä näytetään punaisella ja pari-end lukee sopusoinnussa lineaarisen transkription näytetään sinisellä. (B) samanlainen circRNAs, circular DNAs (eccDNAs) havaitaan perustuu rakenteellisesti lukea variantit Yhdenmukainen circularization tapahtuma kuvattu punaisella .
sen jälkeen, kun ne löydettiin uudelleen, tiedeyhteisö on kiinnittänyt huomionsa circRNAs: iin ja tutkinut niiden osallistumista useisiin soluprosesseihin terveyden ja sairauksien alalla (Haque and Harries, 2017), niiden mahdollista roolia biomarkkereina (Abu and Jamal, 2016) ja niiden sääntelytehtäviä (Floris et al., 2016). Circrnojen tiedetään nyt olevan runsaasti ja stabiileja sytosolissa ja tumassa (Salzman et al., 2012; Jeck ym., 2013; Li et al., 2015)ja on myös todettu vapaaksi biofluideissa (Bahn et al., 2015; Memczak et al., 2015; Chen et al., 2018)ja solunulkoisissa vesikkeleissä (Kyoung Mi et al., 2017). CircRNAs: n biomarkkeripotentiaalia on tutkittu intensiivisesti, itse asiassa on julkaistu monia tapauskontrollitutkimuksia, joissa etsitään eri tavoin ilmaistuja circrnoja, jotka voisivat olla eri sairauksien biomarkkereita. Tähän mennessä circRNAs on yhdistetty useisiin sairauksiin, kuten syöpään (Kristensen et al., 2017; Arnaiz et al., 2018), neurologiset häiriöt (Akhter, 2018), sydän-ja verisuonitaudit (Aufiero et al., 2019)ja immuuniperäiset sairaudet (Iparraguirre et al., 2017; Liu et al., 2019). Samalla niiden biogeneesin, ominaisuuksien, toimintojen ja vaikutusten ymmärtäminen ihmisen biologiassa jää avoimiksi kysymyksiksi alan tutkijoille.
vaikka useimpien piirien toiminta on edelleen tuntematon, on osoitettu, että jotkut piirit voivat toimia mikroRNA-sieninä säätelemällä mikroRNA-tasoja ja niiden toimintaa (Hansen et al., 2013; Memczak et al., 2013; Zheng et al., 2016). Ne osallistuvat geeniekspression säätelyyn säätelemällä parempien geeniensä transkriptiota ja kilpailevat lineaaristen sponging-eli sponging-proteiinien kanssa (Ashwal-Fluss et al., 2014; Li et al., 2018). Mielenkiintoista, ribosomiprofilointitutkimukset ovat viime aikoina osoittaneet, että circRNAs voidaan myös kääntää sekä in vitro että In vivo (Legnini et al., 2017; Yang et al., 2017).
piirien tärkein ominaisuus ja suurin osa niiden erityisominaisuuksista on niiden kiertoluonne. Siksi sen lisäksi, että havaitaan niiden ominainen takaisin-saumattu junction, testaus circlulaarisuus näiden molekyylien, on yksi tärkeimmistä kohdista jokaisessa circRNA tutkimus. Monet tutkimukset ovat kuitenkin perustaneet circrnojen löytämisen kokonaisrna: han ja saattaneet siten tulkita joitakin lineaarisia kimeerisiä transkriptejä circrnoiksi, mikä on johtanut vääriin positiivisiin circRNA-havaintoihin. Tämän ongelman kiertämiseksi useimmat tutkimukset ovat vahvistaneet Total RNA-seq: n löytämien transkriptien sirkulariteetin RNase R: llä, Northern blotilla tai elektroforeettisilla menetelmillä (Jeck and Sharpless, 2014). Kuitenkin nämä circularity validations ovat myös joskus paljastanut transkriptit, jotka näyttävät olevan lineaarinen, sen sijaan, että pyöreä vahvistaa, että havaitseminen circRNAs alkaen total RNA voi johtaa joitakin vääriä positiivisia. Nämä väärät positiiviset ovat johtuneet teknisistä esineistä tai transkriptioista, jotka ovat peräisin epätavallisista tapahtumista, kuten exon-kaksoiskappaleista tai transplicing-tapahtumista (Jeck and Sharpless, 2014; Szabo and Salzman, 2016). Se sanoi, mahdollisuus on löytänyt totta, biologisesti aktiivisia ja toiminnallisia lineaarisia transkriptien jotka sisältävät sekvenssi vastaa backsplicing junction (tästä lähtien kutsutaan chimeric lineaarinen transkriptien), on jossain määrin unohdetaan, koska lähde tällaisen lineaarisen RNA ei ole tunnettu terveitä soluja.
ympyränmuotoinen DNAs Kimeeristen lineaaristen transkriptien lähteenä
suurin osa ihmisen genomista on järjestetty lineaarisiin kromosomeihin, mutta joitakin poikkeuksia on jo pitkään hyväksytty, kuten mitokondriaalinen DNA ja kromosomipoikkeavuudet kuten onkogeenejä kantavat DNA-ympyrät (esim. kaksoisminuutteja) (Benner et al., 1991; Nathanson et al., 2014; Turner et al., 2017)ja rengaskromosomit (Tümer et al., 2004). Vasta viime aikoina eri Pyöreä DNAs kuten microDNAs (Shibata et al., 2012) tai ekstrakromosomaalinen Pyöreä DNAs (eccDNAs) havaittiin myös syntyä suuria osia eri eukaryoottisia genomeja kuten ihmisen ja hiiva (Møller et al., 2015; Kumar et al., 2017; Møller ym., 2018).
ympyränmuotoiset Dnat muodostuvat, kun lineaarisen DNA: n kaksi päätä nivoutuvat yhteen, mikä johtaa ympyränmuotoisen, yleisesti kutsutun murtopisteen risteyksen kaltaiseen risteykseen, joka havaitaan ympyrämuotoisen tapahtuman mukaisten rakenteellisten lukuvarianttien perusteella (Gresham et al., 2010; Møller ym., 2018; Prada-Luengo et al., 2019) (Kuva 1B). Ne vaihtelevat yleensä sadasta emäksestä megabaasipiireihin ja voivat sisältää kokonaisia eksoneja ja geenejä (Shibata et al ., 2012; Møller ym., 2015; Kumar et al., 2017; Turner et al., 2017; Møller ym., 2018) ja vaikka jotkut genomin alueet ovat yleisempiä pyöreässä DNA: ssa (Sinclair and Guarente, 1997;; Møller et al., 2016; Turner et al., 2017; Møller ym., 2018), Useimmat kehämäiset DNA: t näyttävät esiintyvän satunnaisesti (Shibata et al., 2012; Møller ym., 2015; Kumar et al., 2017; Møller ym., 2018).
mielenkiintoista, tuoreessa lehdessä Møller et al. tunnistettu tuhansia eccDNAs leukosyyttien ja lihassolujen terveillä verrokeilla. Ajatuksena tutkia, voitaisiinko eccDNAs transkriboida, mRNA-kirjasto myös sekvensoitiin lihaskudoksesta ja analysoitiin transkriptiotapahtumien varalta havaitun eccDNA: n katkopisteen liitoskohdassa löytäen useita vastaavuuksia (Møller et al., 2018). Tämä havainto viittaa siihen, että pyöreä DNA terveessä kudoksessa transkriboidaan, jolloin syntyy lineaarisia ja polyadenyloituja transkriptejä, jotka kantavat sekvenssin, joka vastaa circrnas: n backsplicing-sekvenssiä (Møller et al., 2018) (Kuva 2).
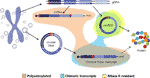
kuva 2 eri transkriptien graafinen esitys joko genomisesta DNA: sta tai kehä-DNA: sta. Eksonit on väritetty purppuralla ja selkäliitokset tai kimeeriset liitokset on esitetty punaisella. Polyadenyloidut, kimeeriset liitokset sisältävät ja RNaasi-R-resistentit transkriptit on korostettu oranssilla, sinisellä ja keltaisella.
ympyränmuotoisten DNAs: ien transkriptiotodisteet yhdessä niiden runsauden kanssa johtavat siihen, että ympyränmuotoiset DNAs: t voisivat olla luonnollinen lähde huomattavalle määrälle lineaarisia RNAS: iä, jotka kuljettavat kimeerisiä liittymiä. Monissa tapauksissa nämä kimeeriset liitokset saattavat olla erottamattomia circrnojen backsplicing-liitoksista, ja siksi ne saattavat olla sekoittavia tekijöitä circRNA-tutkimuksissa. Seuraavissa kohdissa selitämme tätä ehdotusta tukevia tietoja.
sirkrna-detektio: Kaikki, mikä kiiltää, ei ole kultaa
kuten aiemmin esiteltiin, sirkrnat muodostuvat kuitenkin ei-kanonisesta pyörähdystapahtumasta, jota kutsutaan backsplicingiksi. Tästä backsplicing-tapahtumasta johtuvilla transkripteillä on kovalenttisesti suljettu silmukkarakenne, jossa ei ole 5 ’ -3 ’ napaisuutta eikä polyadenyloitua häntää, ja mikä tärkeintä, niille on ominaista se, että ne ovat sekoittuneet eksonijärjestykseen suhteessa lineaariseen transkriptioon (Zhang et al., 2016; Wilusz, 2018). Tämä sekoitettu eksonijärjestys ilmenee backspliced junction, joka yhdistää 5 ’alavirtaan sekvenssi kanssa ylävirtaan 3’ sekvenssi. Siten kaikki circRNA-tunnistusalgoritmit hyödyntävät taustaliitosten läsnäoloa diagnostisena ominaisuutena circRNA: n tunnistamisessa (Kuva 1a).
näiden selkäliitosten havaitsemiseen on sovellettu erilaisia menetelmiä. Kaupallisia ryhmiä, jotka sisältävät luotaimia, jotka on suunnattu näille takaisinkytketyille alueille, on käytetty laajalti biomarkkerien seulontatutkimuksissa (Iparraguirre et al., 2017; Liu et al., 2017; Sui et al., 2017; Li et al., 2018). Myöhempi validointi perustuu usein myös backsplicing-liittymien vahvistamiseen käyttämällä erilaisia alukkeita (Panda and Gorospe, 2018). Monet muut paperit ovat tehneet suuren suoritustehon sekvensointi analyysi, joka voittaa yksi tärkeimmistä rajoituksista taulukoiden avulla havaita paitsi huomautusten circRNAs mutta myös de novo RNA circularization tapahtumia genomialueilta, joissa ei circRNA oli huomautettu aiemmissa tutkimuksissa. Useita bioinformaattisia putkistoja on kehitetty sirkrnojen havaitsemiseen RNA-Seq-tietokokonaisuuksissa, mutta kaikki ne perustuvat siihen, että lukuja on ristikkäin kääntyvien liittymien yli ja luotettavimman löytäminen on edelleen haaste bioinformaatikoille (Hansen et al., 2016; Hansen, 2018; Prada-Luengo et al., 2019).
RNA-Seq-aineistossa voidaan käyttää kahta pääasiallista menetelmää circRNAs-arvojen toteamiseksi. Ensinnäkin monet circRNA-RNA-seq-tutkimukset perustuvat RNaasi-R-käsiteltyihin näytteisiin, jotta kaikki lineaariset Rnat voidaan poistaa ennen sekvensointia. Vaikka tämä lähestymistapa on suunniteltu erityisesti circRNA-havaitsemiseen, on syytä huomata, että RNaasi-R: n hajoaminen on vaihtelevaa, että on olemassa harvinaisia tapauksia RNaasi-R resistenttejä lineaarisia RNAS-ja RNaasi-R-herkkiä circRNAs-tapauksia (Szabo and Salzman, 2016) ja että tämä hoito vaatii suurta RNA-tuloa, joka voisi rajoittaa joidenkin kudosten osalta. Muut circRNA-tutkimukset valitsevat sekvenssin joko kokonais -, ribosomaalinen-köyhdytetty (ribo -), tai ei-polyadenyloitu (polyA -) RNA, jossa sekä lineaarinen ja Pyöreä RNA löytyy (Salzman et al., 2012; Memczak et al., 2013; Broadbent et al., 2015; Lu et al., 2015; Memczak ym., 2015). Tällä lähestymistavalla vältetään RNaasi R: n käyttö, mikä vähentää sekvensointiin tarvittavaa RNA-määrää ja mahdollistaa muiden RNAS-tyyppien ilmentymisen tutkimisen samasta aineistosta. On osoitettu, että hyvällä sekvensointisyvyydellä ja-laadulla ja huolella tehdyllä data-analyysillä voidaan havaita totaalisia sirkrnoja kokonaisrna-sekvensoinnista (Wang et al., 2017), kuitenkin, tässä toisessa lähestymistavassa, tarvitaan myöhemmin circrnas-sirkulariteettivahvistusta.
ympyränmuotoisista Dnoista litteroitujen lineaarikimeeristen Rnojen löydyttyä circrnat eivät ole enää ainoita transkriptejä, joissa on kimeeriset liitokset. Sen vuoksi on erittäin tärkeää huomata, että vaikka ensimmäinen lähestymistapa rikastuttaa RNA-näytettä merkittävästi ympyräpiireissä siten, että suurin osa havaituista kimeerisistä liitoksista vastaa todellisia ympyräpiirejä, toinen menetelmä saattaa yliarvioida circRNA-transkriptien määrän liittämällä circrnoihin signaalin, joka tulee sekä ympyräpiireistä että DNAs-ympyrästä litteroiduista lineaarisista chimeerisistä transkripteistä. Näin ollen, ottaen huomioon rinnakkaiselo circRNAs ja lineaarinen kimeerinen transkriptien, tarve circularity testit ja toiminnallisuus määritykset saa merkitystä ja erityistä varovaisuutta olisi noudatettava ei vain kokeellisia vaan myös laskennallisia menetelmiä, jotta vältetään sekoittaminen kimeerinen transkriptien ympyrän DNAs kanssa circrnas muodostettu backsplicing.
Keskustelu
circRNA-kenttä on vielä alkuvaiheessa, mutta circrnat ovat jo osoittautuneet hämmästyttäviksi molekyyleiksi, jotka ovat sekaantuneet moniin prosesseihin, joilla on suuri biomarkkeripotentiaali ja jotka voivat myös muuttaa tapaa, jolla ymmärrämme transkriptio-ja translaatioprosessit. Näistä syistä ne ovat saamassa huomiota ja circRNA-kenttä on tällä hetkellä yksi aktiivisimmista RNA-tutkimuskentistä. On kuitenkin vielä paljon ristiriitoja, ristiriitoja ja avoimia kysymyksiä (Li, 2019), joista on keskusteltava.
tässä raportissa ja kehämäisen DNA-kentän viimeaikaisten edistysaskelten valossa haluamme huomauttaa ekstrakromosomaalisen kehämäisen DNA: n transkription olevan yksi tärkeimmistä luonnollisista lineaaristen transkriptien lähteistä, joissa on taaksepäin sidottuja signaaleja, jotka voivat häiritä circRNA-tietoja (Møller et al., 2018). Tästä lähtien, teknisten esineiden, kopioiden ja tapahtumien lisäksi, jotka voivat johtaa vääriin positiivisiin tuloksiin circRNA: n havaitsemisessa, meidän olisi otettava huomioon myös tämän uudentyyppisen kimeerisen selostuksen olemassaolo. Siksi kiertokulkutestit ja toiminnalliset määritykset ovat tärkeämpiä kuin koskaan.
joka tapauksessa näitä kimeerisiä lineaarisia transkriptejä ei tulisi pitää pelkästään circRNA-tutkimusten sekoittavina tekijöinä. Huolimatta teknisistä vaikutuksista circRNA-luonnehdintaan, näiden eccdnas: stä tulevien circRNA-kaltaisten kimeeristen lineaaristen RNA-molekyylien olemassaolo lisää uudenlaisen molekyylin alati kasvavaan RNA: n luetteloon ja laajentaa näkemystämme transkriptomin monimutkaisuudesta ja sen säätelystä. Lisäksi näillä eccDNA: sta tulevilla lineaarisilla RNA-molekyyleillä saattoi esiintyä myös circRNA: n kaltaisia funktioita, kuten säätelyfunktioita tai transloituvia mahdollisuuksia. EccDNA-transkriptien geenituotteet voivat mahdollisesti edistää somaattisten solujen ja kudosten fenotyyppiä hiivassa (Gresham et al., 2010; Demeke et al., 2015). Tällä orastavalla alalla tarvitaan kuitenkin lisää tietoa ja tutkimusta, jotta voidaan alkaa raapia jäävuoren pintaa.
tekijän osuudet
LI, IP-L, BR ja DO kirjoittivat paperin.
Rahoitus
tätä tutkimusta on rahoittanut Instituto de Salud Carlos III hankkeella ”Pi17/00189” (Euroopan aluekehitysrahaston/Euroopan sosiaalirahaston yhteisrahoittama) ”Investing in your future”. IP-L: ää ja BR: ää tuki Tanskan riippumaton tutkimusneuvosto, 6108-00171b ja LI: tä tuki Baskimaan hallituksen Opetusministeriö .
eturistiriita
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.