reclame:
In dit artikel zullen we bespreken over het spijsverteringsstelsel van gewervelde dieren met behulp van geschikte diagrammen.
embryonaal spijsverteringskanaal:
Archenteron:
het embryonaal archenteron wordt het slijmvlies van het volwassen spijsverteringskanaal en van alle derivaten daarvan. Splanchnic mesoderm voegt lagen van bindweefsel en gladde spieren rond de archenteron. Ectodermale invaginatie van het hoofd vormt het stomodaeum dat leidt naar de mondholte, en een soortgelijke mid-ventrale ectodermale invaginatie vormt proctodaeum, die leidt naar het achterbeen.
reclame:
het stomodaeum wordt de volwassen buccale holte en geeft aanleiding tot tandglazuur, epitheliale bedekking van tong, klieren, bijvoorbeeld, slijm, gif en speekselholte, enz. en Rathke ‘ s buidel van de hypofyse. Het proctodaeum vormt ofwel een klein einddeel van de cloaca bij lagere gewervelde dieren en het rectum bij zoogdieren.
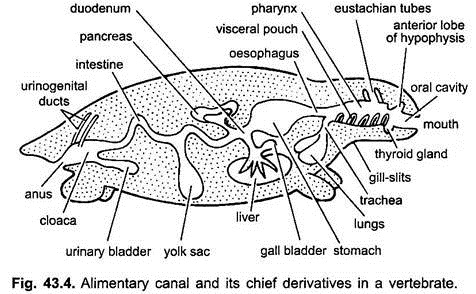
het spijsverteringskanaal bij embryo ‘ s van maag tot cloaca is bevestigd aan de dorsale lichaamswand door een dubbele plooi van buikvlies, het dorsale mesenterium genoemd, en aan de ventrale lichaamswand door een ventraal mesenterium. Bij volwassenen blijft het dorsale mesenterium bestaan, maar het ventrale mesenterium verdwijnt alleen in het gebied van de lever en de urineblaas.
spijsverteringskanaal van volwassenen:
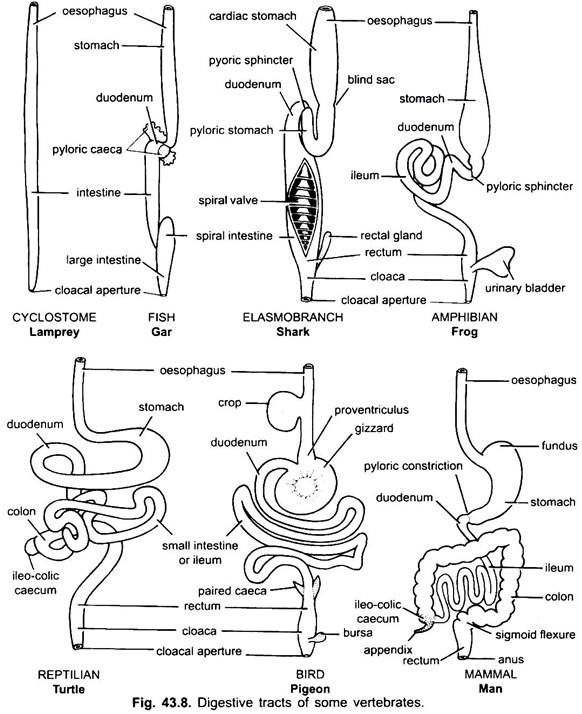
het spijsverteringskanaal onderscheidt zich voor verschillende functies in de volgende gebieden: mond, buccale holte, keelholte, slokdarm, maag, dunne darm, dikke darm en cloaca. Na uitlopers ontstaan uit het spijsverteringskanaal – mondklieren, Rathke ‘ s buidel, schildklier, kieuwspleten, trommelholte, thymus en andere klieren van kieuwspleten, luchtpijp, longen, zwem blaas, lever, pancreas, dooierzak, en urineblaas.
reclame:
histologie:
de wand van het spijsverteringskanaal bestaat uit vier concentrische lagen.
zij zijn:
(i) een buitenste viscerale buikvlies of sereuze vacht bestaat uit mesotheliale cellen en een dunne laag bindweefsel. Het ontbreekt in de slokdarm,
reclame:
(ii) daaronder bevindt zich een spierlaag gevormd door gladde spiervezels die in de buitenlangse en binnencirkelvormige spiervezels zijn aangebracht. Tussen de twee spierlagen bevindt zich een netwerk van zenuwcellen en zenuwvezels van het autonome zenuwstelsel, bekend als myenteric plexus of plexus van Auerbach.
(iii) onder de spierlaag bevindt zich een submucosa gemaakt van bindweefsel met elastische vezels, vet -, bloed-en lymfevaten, zenuwcellen en vezelklieren,
(iv) De binnenste laag is een mucosa bestaande uit drie gebieden:
(a) buitenste meest smalle muscularis mucosa van buitenste longitudinale en binnenste cirkelvormige gladde spiervezels.
(B) middelste dunne laag lamina propria van bindweefsel, bloedvaten, zenuwen en knobbeltjes van lymfatisch weefsel, en
(C) een keldermembraan dat een laag zuilvormige epitheliale cellen ondersteunt die vaak kliervormig en ciliair zijn.
mond:
mond is de opening die naar de buccale holte leidt. Bij lampreys (cyclostomen) is het een cirkelvormige opening aan de basis van de buccale trechter en blijft permanent open door gebrek aan kaken, enz. In gnathostomes is het terminaal. Mond wordt begrensd door de lippen die onbeweeglijk zijn en gevormd van de verharde huid in vissen, amfibieën en reptielen. Bij zoogdieren zijn deze vlezig en gespierd.
buccale holte:
reclame:
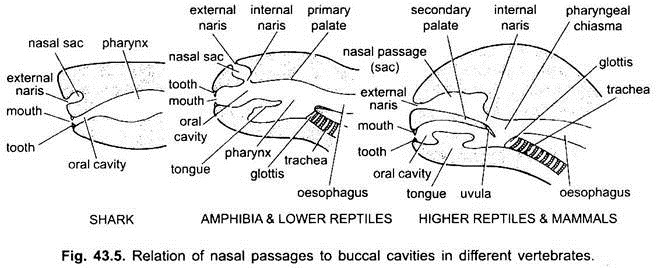
de ruimte tussen de lippen en de kaken is een vestibule. Het kan aan de buitenkant worden begrensd door wangen en aan de binnenkant door het tandvlees. Slijmklieren van Wangen openen zich in de vestibule. De mond opent in een buccale holte, dat is een ruimte tussen de mond en de keelholte. Het exacte punt waar de stomodaeale ectoderm en faryngeale endoderm samensmelten is variabel en niet gemakkelijk te onderscheiden.
bij elasmobranchen en de meeste benige vissen openen de neusholtes zich niet in de buccale holte. Bij Chondrichthyes en tetrapoda (amfibieën en de meeste reptielen) openen de neusholten zich in de buccale holte door choanae of interne nares, die primitief voorin geplaatst zijn, maar bij krokodillen, vogels en zoogdieren worden ze posterieur in de keelholte als gevolg van de vorming van een secundaire gehemelte, die effectief scheidt de luchtwegen neus passage uit de mondholte of voedsel passage.
bij vogels is dit gehemelte gespleten waardoor neus-en buccale holtes met elkaar communiceren. Bij zoogdieren wordt het secundaire gehemelte achteraf voortgezet als een membraneus zacht gehemelte. Bij mensen hangt een zacht gehemelte in de larynx keelholte in de vorm van vlezig proces, huig genaamd.
derivaten van buccale holte:
1. Orale klieren:
er zijn twee soorten integumetaire meercellige klieren die zich in de buccale holte openen. Het zijn slijmklieren en enzymatische klieren. Vissen en aquatische amfibieën hebben alleen slijmklieren. Reptielen hebben klieren in groepen, zoals palatine, linguale, sublinguale en labiale klieren genoemd volgens locatie, ze produceren ook slijm.
bij giftige slangen worden de bovenste labiale klieren aangepast om gif af te scheiden, terwijl in het Gila-monster de sublinguale klieren GIF produceren. Vogels hebben sublinguale klieren en een klier in de mondhoek. Zoogdieren hebben veel kleine slijmklieren naast echte en vergrote speekselklieren die enzymatisch zijn. Ze zijn parotis, sublinguaal, submaxillaire en infraorbitale speekselklieren, afscheiden mucine en ptyaline.
2. Tong:
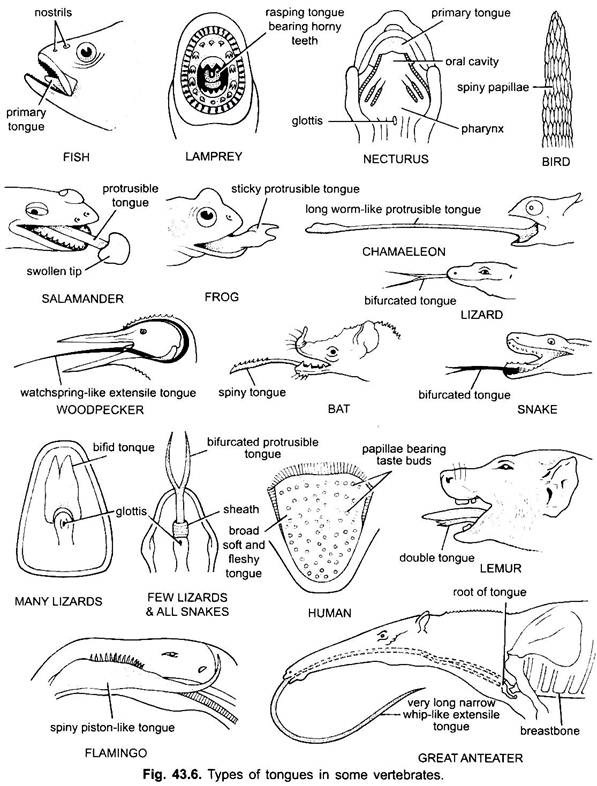
de tong komt voor bij alle gewervelde dieren. Tong in gewervelde dieren vertonen veel diversiteit en zijn niet homoloog. In cyclostomen is er een Gespierde, vlezige, raspende tong met geile tanden om de huid en spieren van hun prooi te raspen.
vissen hebben een primaire tong, gevormd door een vlezige plooi van de buccale vloer. Het heeft geen spieren, maar receptoren en tanden zijn aanwezig op de tong in sommige benige vissen. De tong is bedekt met slijmvlies. Bij sommige amfibieën ontbreekt de tong of is deze onbeweeglijk. De meeste amfibieën, echter, hebben een uitstekende tong en bij sommige kikkers en padden kan het terug gevouwen op zichzelf wanneer niet in gebruik.
het kan door een snelle instroom van lymfe uit de mond worden gegooid om de insecten te vangen. De tong bij hagedissen en slangen is vaak sterk ontwikkeld. In kameleons, is het zeer uitbreidbaar gebruikt om insecten te vangen. De punt is verdikt en plakkerig. De gevorkte punt van de tong bij slangen dient als een middel om chemische stimuli van de externe omgeving naar de gepaarde vomero-nasale organen op het dak van de mond over te brengen.
bij schildpadden en krokodilachtigen kan de tong niet worden uitgeschoven. De vruchtwatertong heeft vrijwillige spieren, het ontvangt de hypoglykemische zenuw en heeft klieren en smaakpapillen. Het ontwikkelt ook intrinsieke spieren die de tong bewegen. Bij vogels is de tong slank en heeft een hoornachtige bekleding. Bij sommige vogels is de tong onbeweeglijk, bij sommige vogels is hij lang, grauw en vaak gebruikt voor het vangen van het voedsel.
bij de meeste zoogdieren, met uitzondering van walvissen, is de tong sterk ontwikkeld en in staat tot aanzienlijke beweging, naast het uitrekken en terugtrekken, als gevolg van de aanwezigheid van een aantal intrinsieke spieren.
bij zoogdieren vormt het slijmvlies onder de tong een mediane plooi, frenulum genaamd, die de tong verbindt met de mondbodem. Bij zoogdieren draagt het bovenoppervlak van de tong vier soorten papillen (filiform, fungiform, foliate en circumvalate), met smaakpapillen behalve filiform papillen.
3. Tanden:
gewervelde dieren hebben twee soorten tanden die aan kaakbeenderen zijn bevestigd: epidermale tanden en echte tanden. Epidermale tanden worden het best ontwikkeld in cyclostomen. Het zijn harde, conische, hoornachtige structuren afgeleid van het stratum corneum. In lampreys zijn ze te vinden op de wanden van de buccale trechter en op de tong. Kikkervisjes larven van kikkers en padden hebben gekartelde epidermale tanden in rijen op de lippen. Bij zoogdieren heeft het volwassen vogelbekdier epidermale tanden.
echte tanden:
tanden worden niet gevonden bij baleinwalvissen en miereneters bij zoogdieren, en agnathanen, steuren, sommige padden, sireniërs, schildpadden en moderne vogels, enz. In lagere gewervelde dieren (zoals vissen, amfibieën en de meeste reptielen) kunnen tanden continu een onbepaald aantal keren worden vervangen, dergelijke tanden worden polyphyodont genoemd. Deze tanden zijn homodont (vergelijkbaar type) en acrodont. (met de kaakbeenderen).
bij de meeste zoogdieren zijn de tanden diphyodont, thecodont en heterodont. Bij sommige zoogdieren zijn dit monofyodont met slechts één set tanden, bijvoorbeeld mollen, Indiase eekhoorn. Tanden zijn qua structuur vergelijkbaar met de placoïde schubben van haaien, gevormd uit een centrale pulpholte, waaromheen een dikke maar zachte laag aanwezig is, de dentine, die extern is bedekt met een dun, extreem hard glazuur. Deze worden verondersteld te zijn afgeleid van benige schubben van ostracodermen en placodermen. Voor details lezers kunnen zien het gebit bij zoogdieren.
4. Adenohypophysis:
de voorste kwab van de hypofyse ontwikkelt zich als een dorsale evaginatie van stomodaeum, genaamd Rathke ‘ s pouch, die zich vernauwt om de voorste en middelste kwab van de hypofyse te vormen (adenohypophysis). De achterste kwab van hypofyse of neurohypophysis is de ventrale evaginatie van diencephalon, genaamd infundibulum. Dus, het is nerveus deel.
keelholte:
het deel van het spijsverteringskanaal direct achter de buccale holte is een keelholte, bekleed met endoderm. Het is een gemeenschappelijke passage die zowel voor de spijsvertering als voor de ademhaling dient. Als onderdeel van het spijsverteringsstelsel wordt het gebruikt als een doorgang voor voedsel van de buccale holte naar de slokdarm, de spieren initiëren slikken.
bij vissen is de keelholte groot en zijdelings geperforeerd voor kieuwspleten, terwijl hij bij tetrapoda kort is en openingen van neusgaten draagt. Bij embryo ‘ s geeft de wand van de keelholte een aantal ontwijkingen af die zich ontwikkelen tot spiracles, kieuwspleten, luchtblazen, longen, amandelen en een paar endocriene klieren (bijv. thymus, schildklier en bijschildklieren).
slokdarm:
bij de meeste vissen en amfibieën is de slokdarm kort omdat de nek ontbreekt, maar bij vruchtwaterpunctie is de slokdarm langer vanwege de aanwezigheid van de nek. De slokdarm van reptielen is langer dan die van vissen en amfibieën. Bij zaadetende en vleesetende vogels wordt een deel van de slokdarm vergroot tot een zakachtig zakje, gewas genaamd, dat dient om snel gegeten voedsel op te slaan.
het gewas heeft in wezen een tekort aan spijsverteringsklieren, hoewel het gewas bij duiven 2 gewas-klieren heeft bij beide geslachten, het zijn eigenlijk geen klieren maar celvormende structuren, de cellen vormen “duivenmelk” die aan de jongen wordt gevoerd. Bij zoogdieren is de slokdarm lang, mist klieren en varieert in verhouding tot de lengte van de nek.
het gaat door het middenrif, het gedeelte onder het middenrif is bedekt met visceraal peritoneum dat ontbreekt aan het bovenste deel. Slokdarm heeft slijmklieren. De bekleding vormt longitudinale plooien, of vinger-achtige vlezige papillen (elasmobranchs) of horny papillen bij zeeschildpadden.
histologisch verschilt de slokdarm op drie punten van de rest van het spijsverteringskanaal:
(i) Het heeft geen visceraal peritoneum omdat het buiten het coelom ligt, de buitenste afdeklaag is een dunne Tunica adventitia.
(ii) de spiervezels in het voorste deel zijn gestreept, het middelste deel heeft zowel gestreepte als niet-gestreepte spieren, en het achterste deel heeft alleen niet-gestreepte spieren. Maar er zijn uitzonderingen bij herkauwende zoogdieren, alle spieren zijn gestreept of vrijwillig.
(iii) het slijmvlies bestaat uit gelaagde plaveiselepitheliale cellen en niet uit zuilvormige cellen.
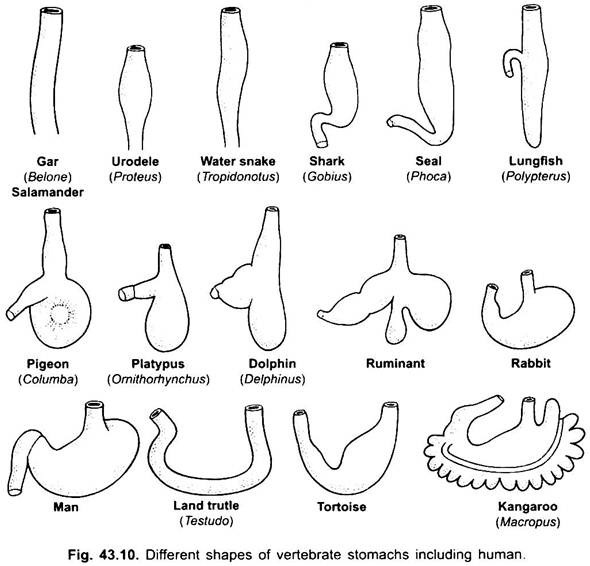
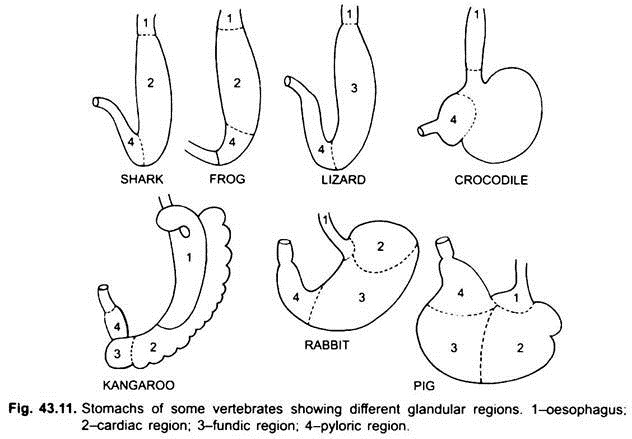
maag:
er is praktisch geen maag in cyclostomen, chimeras, longvissen en sommige primitieve teleostvissen, omdat het geen maagklieren heeft, maar in de meeste vissen en tetrapoda wordt het verwijd voor opslag en maceratie van vast voedsel, en de vertering van voedsel omdat het maagklieren bevat.
het eerste deel van de maag, naast de slokdarm, is het hartstreek en het onderste uiteinde bij de darm is het pylorusgebied, dat een pylorus of pylorusklep heeft, waarin het slijmvlies wordt omgeven door een dikke sluitspier die het openen en sluiten van de pylorus maag in de darm reguleert.
maag is recht in cyclostomen, gar, Belone, enz., en spindelvormig in Proteus, Necturus, sommige hagedissen en slangen. Bij schildpadden en schildpadden is het een brede gebogen buis, en bij elasmobranchs is de maag J-vormig. Bij krokodillen en vogels heeft de maag twee delen, een proventriculus met maagklieren, en een zeer gespierde spiermaag, die het pylorisch gebied vertegenwoordigt en een harde, cornified voering heeft voor het Malen van voedsel.
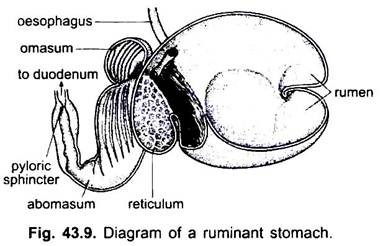
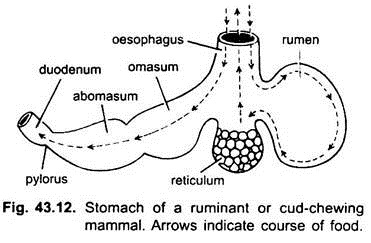
bij zoogdieren ligt de maag dwars en kan het een eenvoudige zak zijn of verdeeld in 3 regio ‘ s, namelijk hart, fundic en pylorisch en elke regio heeft zijn maagklieren. Bij veel herkauwers heeft de maag vier kamers – een pens, reticulum, omasum en een abomasum. Er wordt beweerd dat de eerste drie kamers modificaties van de slokdarm zijn, en abomasum is de ware maag die de hart -, fundische en pylorische delen van de maag vertegenwoordigt. Embryologisch is aangetoond dat alle vier de kamers gemodificeerde gebieden van de maag zijn. Bij kamelen is er geen omasum, de pens en het reticulum hebben pouch-achtige watercellen die ooit werden verondersteld om water op te slaan, maar ze zijn waarschijnlijk spijsvertering.
histologisch heeft de maag de typische delen van het spijsverteringskanaal, maar het heeft twee eigenaardigheden: de muscularis mucosa bestaat uit een buitenste longitudinale laag en een binnenste cirkelvormige spierlaag. Het epitheel voering is dik met verschillende soorten kliercellen vormen maagklieren van drie soorten genoemd hart, fundic, en pyloric maagklieren.
de hart-en pylorusklieren scheiden alleen slijm uit hun oppervlaktecellen af. Fundic klieren (of hartklieren in sommige) hebben drie soorten cellen, slijmhals cellen produceren slijm, oxyntic cellen produceren zoutzuur, ze kunnen ook aanwezig zijn in het hart regio, zymogen cellen of peptic cellen scheiden pepsine.
bij de meeste dieren scheiden de zymogencellen ook twee pro-enzymen af, propepsine en prorennine, die door zoutzuur worden omgezet in respectievelijk pepsine en rennine. De afscheidingen van alle maagcellen vormen een mengsel genaamd maagsap.
Dunne Darm:
de dunne darm is lang, smal en opgerold na de pylorus. Het is het belangrijkste onderdeel van het spijsverteringskanaal omdat de spijsvertering en absorptie van voedsel daarin plaatsvinden. Bij cyclostomen is de darm een korte rechte buis met een spiraalvormig geplaatste longitudinale flap die zich erin uitstrekt.
bij elasmobranches is het verdeeld in kleine en grote porties, en het kleine gedeelte heeft een spiraalklep die het absorberende oppervlak sterk vergroot. Een spiraalklep is ook aanwezig in de dunne darm van een paar meer primitieve benige vissen, maar ontbreekt in hogere vormen waarin de darm lang en opgerold is.
bij caecilians is de maag weinig opgerold en niet onderverdeeld in een klein en groot darmkanaal. Bij kikkers en padden is het relatief lang en opgerold. Bij reptielen is het meer opgerold dan bij amfibieën. Voor het eerst ontstaat bij gewervelde dieren een caecum of blind diverticulum op de kruising van kleine en dikke darm.
dit is echter niet permanent bij alle reptielen. Bij vogels, de dunne darm is opgerold of lus en een of twee koliek caeca zijn ook aanwezig op de kruising van kleine en dikke darm. Bij de meeste zoogdieren is ook de dunne darm proportioneel lang en opgerold. De lengte is echter gecorreleerd met voedselgewoonten. Bij herbivoren is het relatief langer in vergelijking met insectivoren en carnivoren.
er is een blinde zak of caecum op de kruising van de dikke darm en de dunne darm, die meestal klein is bij vleesetende soorten en vrij lang bij veel herbivoren. Het eerste deel van de dunne darm is de twaalfvingerige darm, die kort van pylorus begint en voorbij de ingang van alvleesklier-en leverkanalen eindigt.
het heeft veel gevouwen villi en bevat vertakte Brunnerklieren in de submucosa die slijm, wat alkalisch waterige vloeistof en een beetje enzym afscheiden. Twaalfvingerige darm produceert ook twee hormonen secretine en cholecystokinine die de alvleesklier en galblaas stimuleren om hun sappen te bevrijden. Kanalen van de galblaas en alvleesklier openen in de twaalfvingerige darm.
achter het twaalfvingerige darm bevindt zich een ileum, dat alleen bij zoogdieren wordt onderscheiden in een anterieur kleiner jejunum en later langer ileum. Een groot aantal kleine spijsverteringsklieren zijn aanwezig in de dunne darm. Het zijn buisvormige klieren of crypten van Lieberkuhn gevonden door de hele lengte, ze scheiden slijm en een succus entericus die verschillende enzymen heeft.
het slijmvlies van de dunne darm wordt gevouwen tot kleine villi, die het oppervlak voor secretie en absorptie vergroten. De villi zijn dicht bedekt met minuscule vinger-achtige uitsteeksels, genaamd microvilli die helpen bij de absorptie in de villi. Bij zoogdieren worden knobbeltjes van lymfoïde weefsel genaamd Peyer ‘ s patches gevonden op het ileum.
dikke darm:
dikke darm heeft een grotere diameter dan de dunne darm. Bij vissen, amfibieën, reptielen en vogels is hij over het algemeen kort, maar bij zoogdieren is hij lang. In lagere vormen vormt de dikke darm een rectum, maar in tetrapoda heeft het een dikke darm en terminaal rectum. Bij de meeste vissen en amfibieën leidt het eindgedeelte van het rectum tot een cloaca gevormd door het proctodaeum.
het rectum, de uitscheidingskanalen en de genitale kanalen openen zich in de cloaca en openen zich naar buiten door een cloacale opening. Maar bij veel benige vissen en alle zoogdieren (behalve monotremen) hebben het rectum en de urinewegen aparte openingen aan de buitenkant.; de opening van de eerste is een anus.
het Rectum van zoogdieren is niet homoloog met het rectum van gewervelde dieren, aangezien het bij zoogdieren wordt verkregen door partitionering van embryonale cloaca. In de meeste gewervelde embryo ‘ s is er een postanale darm als een uitbreiding van de darm in de staart, maar het verdwijnt later.
bij elasmobranchen draagt de dikke darm een paar rectale klieren die slijm en natriumchloride afscheiden. In vruchtwaterpunctie is er een ileocolische klep op de kruising van kleine en dikke darmen, die afwezig is in vissen. Het voorkomt dat bacteriën in het ileum van de dikke darm.
bij vruchtwaterpunctie ontstaat een ileocolisch caecum dat bij vogels twee is. Het bevat bacteriën die cellulose verteren. Het is erg lang bij plantenetende zoogdieren (konijn, paard, koe, enz.). Bij primaten is caecum klein met een rudimentair vermiform appendix.
spijsverteringsklieren:
1. Lever:
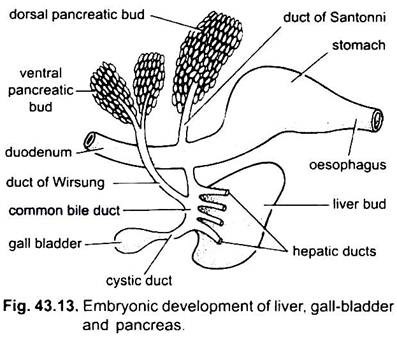
de lever ontstaat als een enkele of dubbele uitgroei uit de ventrale wand van het embryonale archenteron. Deze uitgroei vormt een hol hepatisch diverticulum, dat zich al snel onderscheidt in een voorste deel, dat zich uitbreidt tot de lever en zijn galwegen, en een achterste deel, dat aanleiding geeft tot de galblaas en cystische kanaal. De galwegen verbinden zich tot een hepatisch kanaal dat zich verenigt met het cystische kanaal om een gemeenschappelijk galkanaal of ductus choledochus te vormen. Het gebied van de archenteron waaruit de lever ontstaat wordt de twaalfvingerige darm.
de lever is de grootste gelobde klier in het lichaam, gesuspendeerd door een dubbele laag peritoneum van het transversale septum of zijn vertegenwoordiger.
een galblaas is bedoeld voor de opslag van gal die door de levercellen wordt uitgescheiden, in de lever ligt en in de twaalfvingerige darm afvoert via de gemeenschappelijke galgang die wordt gevormd door de Vereniging van cystische en hepatische kanalen. Een galblaas is niet onmisbaar en ontbreekt bij veel vogels en zoogdieren.
een lever is aanwezig in alle gewervelde dieren. Bij cyclostomen is het klein, enkel gelobd (lampreys) en twee gelobd in hagfishes. Het is gefilobed in elasmobranches, twee of drie gelobd in benige vissen, amfibieën, reptielen en vogels en veel gelobd in zoogdieren. Lever lang, smal en cilindrisch in vissen, urodeles en slangen.
het is kort, breed en afgeplat bij vogels en zoogdieren. Een galblaas en galkanaal zijn aanwezig in larvale cyclostomen, maar ze zijn afwezig in de volwassen. Vissen, amfibieën en reptielen hebben over het algemeen een galblaas, maar het ontbreekt bij veel vogels. De meeste zoogdieren hebben een galblaas, maar deze is afwezig in Cetacea en Ungulata.
de lever scheidt een waterige, alkalische gal af, maar heeft geen enzymen. Het neutraliseert de zuurgraad van voedsel dat de twaalfvingerige darm binnenkomt. Het helpt bij de vertering van vetten.
2. Pancreas:
Pancreas wordt gevormd uit de endoderm van het embryonale archenteron. Een enkel dorsaal diverticulum uit embryonale twaalfvingerige darm en een of twee ventrale uitlopers uit de lever vormen pancreas diverticula. De proximale delen van diverticula vormen pancreas kanalen, maar deze kanalen ondergaan vermindering of fusie, zodat slechts één of twee pancreas kanalen blijven in de VOLWASSENE, ze openen in de twaalfvingerige hetzij afzonderlijk of na vereniging met de gemeenschappelijke galgang.
de distale delen van diverticula ondergaan ontknoping om de hoofdmassa van pancreascellen te vormen waaraan mesodermale derivaten worden toegevoegd. Zo wordt een enkele klier gemaakt die verscheidene lobben heeft die of een diffuse of een compacte alvleesklier vormen.
de alvleesklier is zowel een exocriene als endocriene klier, aan elkaar gebonden door delicate bindweefselstrengen. Het exocriene deel scheidt spijsverteringsenzymen af die in de twaalfvingerige darm door alvleesklierkanalen worden gegoten. Terwijl het endocriene deel hormonen zoals insuline en glucagon afscheidt.
bij alle gewervelde dieren is een alvleesklier aanwezig. Bij lampreys, sommige benige vissen, longvissen en lagere tetrapoden, is het een diffuus orgaan ingebed in de lever, mesenteries en darmwand. Hagfishes hebben een kleine alvleesklier. Elasmobranchs hebben een goed gedefinieerde bilobed pancreas. Bij hogere tetrapoda is het meestal een compacte klier. Een of twee alvleesklier kanalen openen in de twaalfvingerige darm.