- Inleiding
- resultaten
- de DBH-Cre Driver Marks en is beperkt tot de TH-expressie noradrenerge regio ‘ s in de hersenstam die worden geremd door CNO toediening
- CNO-hM4D gemedieerde verstoring van noradrenerge neuronen bij volwassen muizen
- CNO-hM4D verstoring van DBH-Cre neuronen resulteert in een Hypercapnisch tekort
- vaste Broer / Zus controles geïnjecteerd met 10 mg/kg CNO vertoonden een Hypercapnisch Ventilatiedeficit
- Gewend en Unhabituated Wildtype Dieren Ingespoten Met een Zoutoplossing Toonde Geen Wijzigingen Presaline en Postsaline
- Clozapine-CNO-Concentratieratio ‘ s zijn hoger in de hersenen dan in Serum
- discussie
- materialen en methoden
- ethische goedkeuring
- fokken, genetische achtergrond, en onderhoud van muizen
- histologie
- elektrofysiologie
- plethysmografie
- vloeistofchromatografie-massaspectrometrie
- gegevensverzameling en-analyse
- plethysmografie
- statistieken
- plethysmografie
- vloeistofchromatografie-massaspectrometrie
- ethische verklaring
- Auteursbijdragen
- financiering
- belangenverstrengeling verklaring
- Dankbetuigingen
Inleiding
de neurale netwerken die ten grondslag liggen aan de respiratoire chemosensorische reflex zijn een primair doel voor het begrijpen van de etiologie van verschillende gedrags-en fysiologische stoornissen. Verstoorde chemosensorische reflexen worden verondersteld om een rol te spelen in zowel aangeboren als volwassen aandoeningen, waaronder wiegendood syndroom (SIDS), aangeboren Centrale hypoventilatie syndroom (CCHS), centrale slaapapneu ‘ s, abnormale ademhaling in Rett syndroom, en obesitas hypoventilatie syndroom (Obonai et al., 1998; Ozawa et al., 2003; Amiel et al., 2009; Ramanantsoa et al., 2011; Lavezzi et al., 2013). Onder de valse verstikkingsalarmhypothese wordt verondersteld dat ongepaste chemosensorische activering of overgevoeligheid een rol spelen bij subgroepen van patiënten die lijden aan paniekstoornis (Klein, 1993). Chemosensorische disfunctie kan ook een rol spelen bij neurodegeneratieve ziekten door slaap-wanordelijke ademhaling die wordt geassocieerd met versnelde progressie (Hakim et al., 2012; Verbraecken and McNicholas, 2013; Bahia and Pereira, 2015; Snyder et al., 2017). Zo zal een beter begrip van hersenstam chemosensorische netwerken belangrijke aanwijzingen voor een aantal gedrags-en fysiologische pathologieën.
Pharmacogenetic designer receptors exclusively activated by designer drugs (DREADDs) (Armbruster et al., 2007) zijn gebruikt in een aantal studies om neurale populaties in de respiratoire chemoreflex in kaart te brengen (ademhaling reactie op verhoogde bloed CO2 niveaus). DREADD-technologie in combinatie met intersectionele genetische implementatie is gebruikt om zeer gerichte neuronale populaties stil te leggen terwijl het observeren van de ademhalingsfunctie in bewuste en ongeremde muizen door ons lab en anderen, het vermijden van veel verwarring van eerdere circuit mapping benaderingen (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Deze en de meeste andere studies omvatten bijna altijd CNO alleen niet-DREADD expressie broer of zus controles die geen chemosensorische of andere respiratoire effecten vertoonden, met het argument dat CNO had geen off-target effecten op de ademhaling bij bewuste en ongeremde muizen ondanks de hoge dosis gebruikt, 10 mg/kg. Niettemin, werd het gevonden in verscheidene andere studies dat CNO en zijn producten van het terugmetabolisme off-target gevolgen op gedrag en beweging in een verscheidenheid van analyses konden hebben, maar ademhalingsoutput werd niet gericht (Guettier et al., 2009; Joober, 2010; MacLaren et al., 2016; Gomez et al., 2017; Ilg et al., 2018; Mahler and Aston-Jones, 2018; Manvich et al., 2018; Padovan-Hernandez en Knackstedt, 2018). Er werd ook aangetoond dat CNO en zijn metabolieten niet gelijkwaardig worden verdeeld tussen de bloedsomloop en de hersenen (Gomez et al., 2017).
een tweede, niet-aangepakt probleem vloeit voort uit de stress die wordt veroorzaakt door het experimentele paradigma, waaronder het houden van dieren in een barometrische plethysmografie kamer, hantering, intra-peritoneale injectie en rectale Temperatuurmetingen. In eerdere dreadd respiratoire studies, werden naïeve muizen geïntroduceerd in een kleine kamer (140-400 mls) en toegestaan om te acclimatiseren 20-40 minuten voor data-acquisitie (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Het is echter niet duidelijk of deze tijd voldoende is om stress-geïnduceerde ademhalingsveranderingen die kunnen werken als een interagerende factor met de effecten van clozapine tot een minimum te beperken.
In onze studies naar de rol van het noradrenerge (NA) systeem in de ademhalingscontrole, trachtten we gebruik te maken van de gevestigde RC::P_hM4D DREADD allel (Ray et al., 2011) om de rol van na-neuronen in baseline en hypercapnische ademhaling in ongebreidelde volwassen dieren te testen met behulp van volledige lichaam plethysmografie. Omdat noradrenaline en NA-neuronen ook een centrale rol spelen in stressreacties (Valentino en van Bockstaele, 2008; Chen et al., 2018), hebben we een reeks studies uitgevoerd om uitgebreide gewenning (meervoudige blootstelling aan het experimentele paradigma waarbij het dier leert dat de ervaring niet bedreigend of overleefbaar is) en hoog vs. lage (1 mg/kg) CNO doses met eerder gepubliceerde DREADD protocollen (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Hier laten we voor het eerst zien dat hoge systemische doses CNO in staat zijn om off-target effecten op een autonome respiratoire functie in bewuste muizen uit te lokken. We onthullen ook dat het off-target effect van CNO op chemosensorische respiratoire output effectief wordt ontmaskerd door uitgebreide gewenning en daarom niet duidelijk zou zijn in eerdere CNO-controlestudies die geen gewenning hadden bij dieren voorafgaand aan de respiratoire meting, waarbij slechts een korte acclimatiseringsperiode werd gebruikt voorafgaand aan het verzamelen van gegevens. Samen, stellen deze resultaten voor dat eerder in kaart gebrachte neuronenpopulaties indirect ademhalingscontrole door potentiële rollen in het regelen van spanningsreacties kunnen beà nvloeden. Deze gegevens stemmen met name overeen met recente rapporten die suggereren dat CNO bij hoge doses niet biologisch inert is via metabole omzetting in clozapine (MacLaren et al., 2016; Gomez et al., 2017) en dat off-target gedragseffecten kunnen manifesteren niet alleen van verstoring van gedragscircuits, maar ook van verstoringen aan onderliggende autonome circuits en homeostase.
resultaten
de DBH-Cre Driver Marks en is beperkt tot de TH-expressie noradrenerge regio ‘ s in de hersenstam die worden geremd door CNO toediening
om de expressie en specificiteit van de DBH-Cre lijn te onderzoeken, gebruikten we een enkel recombinase fokschema (figuur 1A) waarbij we de driver overstaken naar de AI9 lijn (Madisen et al., 2010), die een floxed tdTomato uitdrukt. Kleuring met een tyrosinehydroxylase (TH) antilichaam toonde aan dat de expressie van tdTomato in de hersenstam beperkt was tot TH-expressiegebieden, waaronder de locus coeruleus, A5, A1, A2, A7, SubCV en SubCD kernen zoals verwacht (figuur 1B). Om te bevestigen dat na-neuronen die de hm4d-receptor uitdrukken reageren op CNO, hebben we opnames uitgevoerd op locus coeruleus (LC) – neuronen, waar we een remming van neuronen die vuren bij CNO-badtoepassingen waargenomen (n = 3, figuur 1C).
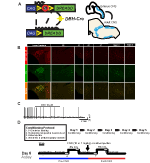
figuur 1. Fokschema en ademhalingsprotocollen. De RC::P_hM4D-allel wordt gecombineerd met een dopamine-beta-hydroxylase (DBH)-Cre-lijn om celspecifieke expressie van de hm4d-receptor in noradrenerge (NA) neuronen (A) te bereiken. Fluorescerende expressie van tdTomato in DBH-Cre; AI9 coronale hersensecties komen samen met tyrosine-hydroxylase (TH) in hersenstam-na-kernen (B) tot expressie. In slice registratie van na locus coeruleus voor en na CNO toepassing, demonstreren DREADD receptor functionaliteit in gerichte neuronen (C). Ademhalingsprotocol. Habituated muizen worden onderworpen aan een 5-daagse gewoonte protocol bestaande uit behandeling, rectale sonde temperatuurmeting, zoutoplossing injectie, en blootstelling aan de plethysmography kamer. Op de zesde dag, zowel habituated als unhabituated muizen onder een hypercapnic assay waar het dier wordt geplaatst in de ademhalingskamer en toegestaan om te acclimatiseren onder basislijn kamer lucht voorwaarden. Het dier wordt vervolgens blootgesteld aan 20 min van 5% CO2, gevolgd door 20 min lucht in de ruimte. Het dier wordt vervolgens intraperitoneaal geïnjecteerd met CNO of zoutoplossing, gevolgd door nog eens 20 min na injectie kamerlucht, 20 min van 5% CO2 en 20 min van kamerlucht. Open vakken omlijnen data verzamelen tijden (D).
CNO-hM4D gemedieerde verstoring van noradrenerge neuronen bij volwassen muizen
om de rol van NA-neuronen onder baseline en hypercapnische ademhaling te onderzoeken, gebruikten we het RC:P_hM4D inhibitory DREADD systeem gekruist met de DBH-Cre driver. Met behulp van het hele lichaam plethysmografie (Ray et al., 2011), hebben we de beademingsreacties gemeten van ongeremde volwassen dieren onder kamerlucht (21% O2/79% N2) en hypercapnische (5% CO2/21% O2/74% N2) condities voor en na CNO toediening (figuur 1D). Om de CNO-dosering en de potentiële stress die door onze experimentele opzet werd veroorzaakt, aan te pakken, werden dieren onderworpen aan een van de vier voorwaarden: (1) niet gehabitueerd en geïnjecteerd met 10 mg/kg CNO; (2) gewend en geïnjecteerd met 10 mg/kg CNO; (3) niet gehabitueerd en geïnjecteerd met 1 mg/kg CNO; of (4) gewend en geïnjecteerd met 1 mg/kg CNO. De gewenning bestond uit een 5-daagse procedure met hantering, rectale temperatuursonde, zoutoplossing injectie en blootstelling aan de plethysmografie kamer elke dag gedurende 30 minuten, terwijl naïeve dieren werden alleen blootgesteld aan een 20-45 minuten kamer acclimatatie periode onmiddellijk voor het verzamelen van gegevens, zoals gedaan in eerdere studies (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). De gemeten respiratoire parameters omvatten respiratoire RR, getijdenvolume (VT), minutenventilatie (VE), zuurstofverbruik (VO2), minutenventilatie genormaliseerd naar zuurstofverbruik (VE/VO2), apneu-frequentie, zuchtfrequentie en variatiecoëfficiënten voor interbreathinterval en amplitude (periodieke en volumeinstabiliteit). Als extra controle, vergeleken we ook gewennde en niet-gehabiteerde wildtype dieren geïnjecteerd met zoutoplossing.
CNO-hM4D verstoring van DBH-Cre neuronen resulteert in een Hypercapnisch tekort
onder drie van de Voorwaarden, DBH-Cre; RC::P_hm4d-dieren vertoonden een afname in VE en VE / VO2 na toediening van CNO, terwijl de broer / zus-controles geen verschillen vertoonden in pre – en post-CNO-waarden. Niet-gehabiteerde dieren die werden geïnjecteerd met 10 mg / kg CNO vertoonden een significant verminderde RR (-12,17%, p = 0,034) en VT (-30,87%, p = 0,0016), resulterend in een afname in VE (-38,64%, p = 0,0031) en een lichte afname in VO2 (-14,25%, p = 0,042) (Figuur 2). De afname in VE was groter dan de afname in VO2, resulterend in een totale afname in VE/VO2 (-26,89%, p = 0,0095). Unhabituated dieren ingespoten met 1 mg/kg van CNO toonde een trend naar kleinere RR (-12.88%, p = 0.066) en een aanzienlijk gereduceerde VT (-16.52%, p = 0.00085) en VIJF (-28.08%, p = 0.0070), wat leidt tot een algemene vermindering in VE/VO2 (-22.23%, p = 0.016) (Figuur 3). Ten slotte vertoonden de dieren die werden geïnjecteerd met 1 mg/kg CNO een verminderde RR (-10,77%, p = 0,074), een trend in de richting van een verminderde VE (-25,07%, p = 0,074) en een verminderde totale VE/VO2 (-23,70%, P = 0,024) (Figuur 4). Geen significante veranderingen in apneu frequentie, zucht frequentie, periodieke of volume instabiliteit, of temperatuur werden gezien in een cohort.
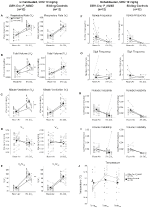
Figuur 2. Niet-gehabiteerde DBH-Cre; RC::P_hM4D dieren die 10 mg/kg CNO toegediend kregen vertonen een hypercapnisch tekort, terwijl de controle van de broer of zus geen verandering vertoont. Na CNO-toediening, DBH-Cre; RC::P_hm4d-dieren vertonen geen verandering in de ventilatie van de lucht in de kamer en verminderingen van RR (A), VT (B), VE (C), VO2 (D) en VE/VO2 (E) Onder hypercapnische omstandigheden zonder veranderingen in de apneu-frequentie (F), zuchtfrequentie (G), periodieke (H) of volumeinstabiliteit (I) en temperatuur (J). Sibling controles toonden geen verschil in alle pre – en post-CNO waarden.
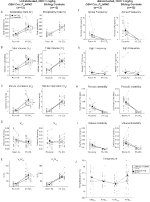
Figuur 3. Niet-gehabiteerde DBH-Cre; RC::P_hM4D dieren die 1 mg/kg CNO toegediend kregen vertonen een hypercapnisch tekort, terwijl de controle van de broer of zus geen verandering vertoont. Na CNO-toediening, DBH-Cre; RC::P_hm4d-dieren vertonen geen significante verandering in de ventilatie van de lucht in de kamer of RR (A) en vertoonden verminderingen in VT (B), VE (C) en VE/VO2 (E) Onder hypercapnische omstandigheden zonder veranderingen in VO2 (D), apneu-frequentie (F), zuchtfrequentie (G), periodieke (H) of volumeinstabiliteit (I) en temperatuur (J). Sibling controles toonden geen verschil in alle pre – en post-CNO waarden.
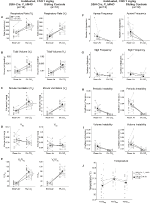
Figuur 4. Gewone DBH-Cre; RC::P_hM4D dieren die 1 mg/kg CNO toegediend kregen vertonen een hypercapnisch tekort, terwijl de controle van de broer of zus geen verandering vertoont. Na toediening van CNO, DBH-Cre; RC:: P_hM4D dieren vertonen geen verandering in de ventilatie van de lucht in de kamer en een vermindering van RR (A) en VE/VO2 (E) Onder hypercapnische omstandigheden zonder significante veranderingen in VT (B),VE (C),VO2 (D), apneu frequentie (F), zucht frequentie (G), periodieke (H) of volumeinstabiliteit (I), en temperatuur (J). Sibling controles toonden geen verschil in alle pre – en post-CNO waarden.
vaste Broer / Zus controles geïnjecteerd met 10 mg/kg CNO vertoonden een Hypercapnisch Ventilatiedeficit
in beide DBH-Cre; RC::P_hm4d en broer of zus controle gewuikte dieren geïnjecteerd met 10 mg/kg CNO, merkten we een significante vermindering van VE / VO2 (p = 0,0235) gemedieerd door dalingen in RR (p = 0,00036) en VE (p = 0,037) (Figuur 5). In tegenstelling tot de andere cohorten was er echter geen verschil tussen DBH-Cre; RC::P_hM4D en sibling controledieren in deze parameters: RR (-12,72 vs.-10,22%, p = 0,6268), VE (-24,88 vs. -23,94%, p = 0,4150), of VE/VO2 (-15,4 vs. -22,55%, p = 0,4643).
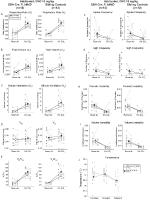
Figuur 5. Beide gewend DBH-Cre; RC::P_hM4D dieren en broer/zus controles toegediend 10 mg / kg CNO vertonen een hypercapnisch tekort. Na toediening van CNO vertonen DBH-Cre; RC::P_hM4D-dieren en controleorganen van broers en zussen geen verandering in de ventilatie van de kamerlucht en een vermindering van RR (A), VE (C) en VE/VO2 (E) Onder hypercapnische omstandigheden zonder significante veranderingen in VT (B), VO2 (D), apneu-frequentie (F), zuchtfrequentie (G), periodieke (H) of volumeinstabiliteit (I) en temperatuur (J).
Gewend en Unhabituated Wildtype Dieren Ingespoten Met een Zoutoplossing Toonde Geen Wijzigingen Presaline en Postsaline
Hoewel er geen fenotypen werden waargenomen in de besturingselementen geïnjecteerd met 1 mg/kg van CNO, we ingegaan op de mogelijkheid dat de injectie zelf veroorzaakt het fenotype waargenomen in de besturingselementen geïnjecteerd met 10 mg/kg van CNO door het testen van wennen en unhabituated wildtype dieren ingespoten met een zoutoplossing (Figuur 6). In zowel gewennde als niet-gehabiteerde cohorten vertoonden de dieren geen verschil in respiratoire parameters presaline en postsaline toediening. Geen significante veranderingen in apneu frequentie, zucht frequentie, periodieke of volume instabiliteit, of temperatuur werden gezien.
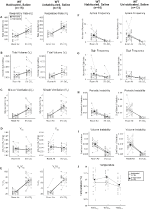
Figuur 6. Zowel niet-gehabiteerde als gewuikte wildtype dieren die zoutoplossing toegediend kregen, vertonen geen verschil tussen de waarden voor en na de injectie. Na toediening van een zoutoplossing vertoonden zowel niet-gehabiteerde als gewuikte wildtype-dieren geen veranderingen onder lucht-of hypercapnische omstandigheden in RR (A), VT (B), VE (C), VO2 (D), VE/VO2 (E), apneu-frequentie (F), zuchtfrequentie (G), periodieke (H) of volumeinstabiliteit (I) en temperatuur (J).
Clozapine-CNO-Concentratieratio ‘ s zijn hoger in de hersenen dan in Serum
om de biologische beschikbaarheid van CNO en clozapine te bepalen, hebben we hun concentraties in serum en hersenen gemeten via massaspectrometrie. Dertig minuten na een Intraperitoneale injectie van CNO bij muizen wordt CNO gevonden in een lagere relatieve abundantie in vergelijking met zijn backmetaboliet clozapine in het serum en de hersenen voor alle geteste doses, 0,1 mg/kg (serum p = 0,0054, hersenen p = 0,0001) (figuur 7A), 1 mg/kg (serum p > 0,05, hersenen p = 0,0197) (figuur 7B) en 10 mg/kg (serum p = 0,0036, hersenen p = 0.0005) (figuur 7C). Wanneer de totale clozapine – CNO-ratio ‘ s werden geanalyseerd, waren ze altijd boven nul en waren ze hoger in de hersenen dan in het serum (vehiculum p > 0,5, 0,1 mg/kg p > 0,5, 1 mg/kg p = 0,0018, 10 mg/kg p = 0,0160) (figuur 7D).
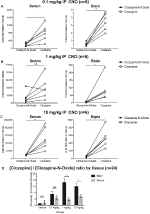
Figuur 7. De concentratieratio ‘ s Clozapine en CNO zijn hoger in de hersenen dan in het serum. Serum-en hersenmonsters werden verzameld bij wildtype muizen 30 min na toediening van CNO in 3 verschillende doses 0,1 mg/kg (A), 1 mg/kg (B), 10 mg / kg (C). Individuele gegevenspunten vertegenwoordigen de relatieve abundantie van clozapine-N-oxide of clozapine verkregen door LC-MS Clozapine tot CNO-ratio voor elke dosis en SEM worden ook getoond (D).
discussie
het oorspronkelijke doel van deze studie was het onderzoeken van de rol van DBH-Cre neuronen in de ademhalingsfysiologie na acute verstoring bij volwassen dieren zonder beperking en bij bewustzijn. Omdat noradrenaline en NA-neuronen een goed gedocumenteerde rol spelen in stressgedrag (Valentino and van Bockstaele, 2008; Chen et al., 2018), probeerden we ook te onderzoeken of gewenning aan een waarschijnlijk stressvol fysiologisch protocol een effect zou hebben op de eerder waargenomen respiratoire fenotypen. In het hele lichaam plethysmografie DREADD protocol gebruikt in ons lab en anderen, dieren worden behandeld, rectaal onderzocht voor temperatuur, blootgesteld aan een nieuwe omgeving (de plethysmografie kamer), en intraperitoneaal geïnjecteerd. Eerdere studies hebben aangetoond dat behandeling en gewenning aan injectie en andere “routine” procedures gedragsmatige en fysiologische parameters, waaronder ademhaling kunnen wijzigen (Misslin et al., 1982; Andrews and File, 1993; Lapin, 1995; Ryabinin et al., 1999). Andere toegepaste stressoren wijzigen ook de ademhaling onder zowel baseline-als hypercapnische ventilatieomstandigheden (isom en Elshowihy, 1982; Kinkead et al., 2001).
in onze studies waarbij hoge doses CNO werden gebruikt, vonden we dat hm4d-gemedieerde remming van DBH-Cre gedefinieerde na-neuronen resulteerde in een verminderde hypercapnische reflex in VE/VO2 in vier gedefinieerde experimentele cohorten, met verminderde RR, VT en VE, ter ondersteuning van eerdere studies (Biancardi et al., 2008; Viemari, 2008; Gargaglioni et al., 2010). In eerdere respiratoire studies werd echter een beademingsdeficit onder hypercapnische omstandigheden aangetoond van dezelfde omvang als bij DBH-Cre; RC::P_hM4D dieren. Geen andere broer of zus controlegroepen vertoonden dit fenotype, inclusief het gewuikte cohort dat alleen (1 mg/kg CNO) of zoutoplossing kreeg. Deze resultaten suggereren dat hogere CNO-doses (10 mg/kg) een effect hebben op de hypercapnische respons die wordt ontmaskerd na uitgebreide gewenning, terwijl ze vermoedelijk de stressniveaus bij dieren verminderen, en dat lagere CNO-doses geen effect hebben op de ademhalingscontrole bij gewennende dieren. Deze resultaten zijn ook in overeenstemming met het werk van Korsak et al. de who heeft aangetoond dat een lage dosis CNO (2 mg / kg) Geen off-target effecten in de werkcapaciteit veroorzaakt in een inspanningstest die voorafgaande training omvatte (Korsak et al., 2018) en Fleury Curado et al. de who toonde geen lage dosis CNO (1 mg/kg) specifieke effecten op de respiratoire output (Fleury Curado et al., 2018).
de verhoogde ratio ‘ s van clozapine en CNO-spiegels in serum en in de hersenen (Figuur 7) stemmen overeen met recente studies die erop wijzen dat CNO gemakkelijk terug gemetaboliseerd wordt tot clozapine en een grotere permeabiliteit van de hersenen vertoont in vergelijking met CNO bij muizen en elders (Jann et al., 1994; Chang et al., 1998; Guettier et al., 2009; Gomez et al., 2017; Raper et al., 2017). Het is echter niet duidelijk of de waargenomen off-target effecten te wijten zijn aan CNO of clozapine. Aangezien onze hypercapnische metingen <30 min na toediening van CNO optraden, is het waarschijnlijk dat de buiten het doel liggende ademhalingseffecten worden gemedieerd door clozapine. Onze resultaten (Figuur 7) tonen hoge relatieve niveaus van clozapine in de hersenen, hoewel CNO niet volledig afwezig is. Huckstepp en collega ‘ s gebruikten echter directe CNO-toepassing op het ventrale medulla in verdoofde ratten om aan te tonen dat alleen bij kamerlucht, en niet onder hypercapnische of hypoxische uitdagingen, CNO-toepassing een klein effect heeft, de frequentie verhoogt en de expiratoire duur vermindert, maar VT onveranderd laat, zonder duidelijk effect waargenomen tijdens hypercapnia (huckstepp et al., 2015). Gezien de directe toepassing bij verdoofde ratten en het tijdsbestek van experimenten, is het waarschijnlijk dat de kleine buiten het doel waargenomen effecten werden gemedieerd door CNO en niet door clozapine.
de backmetaboliet clozapine is een veel gebruikt sedatief en antipsychoticum bij schizofrenie met veel endogene doelwitten, waaronder antagonistische effecten met een lage hoge affiniteit op onder andere D1 -, D2-en D4-dopaminerge receptoren, 5ht2a -, 5HT2C -, 5HT3 -, 5HT6-en 5ht7-serotonerge receptoren, H1-histaminereceptoren en α1-en α2-adrenerge receptoren (Fitton en Heel, 1990; Ashby en Wang, 1996). De hier waargenomen effecten buiten het doel kunnen het gevolg zijn van verschillende of gecombineerde mechanismen en doelstellingen. Clozapine kan de ademhaling als sedatief beïnvloeden. Een eerdere studie toonde verminderde RR en VT Onder 5 en 10% CO2 tijdens zowel langzame-Golf-slaap en snelle-oog-beweging slaap toestanden in vergelijking met de Stille wakker toestand bij muizen (Nakamura et al., 2007). Als alternatief kan remming van gerichte DREADD-expressie neuronen resulteren in een antianxiogeen of anxiolytisch effect vergelijkbaar met ons gewenningsprotocol om CNO/clozapine-gemedieerde chemosensorische off-target effecten te onthullen. Beide verklaringen worden ondersteund door verschillende studies die hebben aangetoond dat CO2 een rol speelt in aangeboren en aangeleerde angstreacties en angstgerelateerd gedrag (Ziemann et al., 2009; Feinstein et al., 2013; Taugher et al., 2014; Dlouhy et al., 2015; Winter et al., 2017). Dus, neuronen gericht in sommige van deze studies kunnen inderdaad een rol spelen in het besturen van anxiogene gedragsreacties in plaats van fysiologische chemosensorische reflexen als zowel catecholaminerge en serotonerge systemen zijn betrokken bij angst / angst gedrag en chemosensorische homeostase (Brust et al., 2014; Hennessy et al., 2017).
omgekeerd kunnen de chemosensorische fenotypen waargenomen met hoge CNO-spiegels echt zijn, aangezien we in staat waren het na-gemedieerde hypercapnisch tekort te recapituleren bij CNO-doses, een orde van grootte lager bij gewuikte muizen, terwijl de controlegroepen geen CNO-effect vertoonden. Volledige vergelijkingen tussen eerdere studies zijn echter moeilijk vanwege het ontbreken, in sommige gevallen, van gerapporteerde VO2 -, VT -, RR-en VE/VO2-gegevens. Veranderingen in lichaamstemperatuur of stofwisseling kunnen bijvoorbeeld ook op een aantal manieren van invloed zijn op de luchtweg-en chemosensorische output, en dat de temperatuur van de plethysmograafkamer in sommige gevallen sterk verschillend was (34 vs.30°C in onze studies), wat het dynamische bereik van de barometrische component van de golfvorm en dus het getijdenvolume beïnvloedt. Met name zagen we noch merkbare veranderingen in respiratoire golfvorm kenmerken in een van onze voorwaarden noch acute cardio-respiratoire arrestatie die werd waargenomen in onze eerdere hoge dosis, hele rhombomere studies (Sun et al., 2017).
onze resultaten tonen voor het eerst aan dat CNO een onverwacht effect heeft op de hypercapnische chemosensorische reflex die wordt ontmaskerd door uitgebreide gewenning. Belangrijk, ondanks hoge niveaus, een CNO off-target effect was eerder uitgesloten vanwege het gebrek aan fenotype in broer of zus controles, maar die we laten zien wordt duidelijk bij gewenning. We bieden een off-target karakterisatie van CNO in het muismodel systeem ter aanvulling van studies bij ratten en niet-menselijke primaten. De resultaten hier verhogen de mogelijkheid dat er extra CNO-gemedieerde, off-target effecten op de onderzochte circuits of op autonome of homeostatische circuits kunnen bestaan, maar kunnen worden gemaskeerd in andere gecontroleerde experimenten. Belangrijk, onthullen deze gegevens dat de onderzoekers zouden moeten streven om de minimale dosissen van activerende ligand mogelijk in combinatie met hoge niveaus van gewenning te gebruiken, en dat de juiste controles in chemische genetische manipulaties moeten worden opgenomen om experimentele gegevens volledig te waarderen en te interpreteren.
materialen en methoden
ethische goedkeuring
Studies werden goedgekeurd door het Baylor College Of Medicine Institutional Animal Care and Use Committee onder protocol AN-6171.
fokken, genetische achtergrond, en onderhoud van muizen
we hielden kolonies van al onze heterozygote muizenstammen in stand door backcrossing naar c57bl/6J muizen en homozygote stammen door broederlijke kruisingen. Voor histologische experimenten werden DBH-Cremice gedekt met de homozygote AI9 muis (Madisen et al., 2010) (JAX 007909). Voor plethysmografie experimenten, DBH-Cre muizen werden gedekt met homozygote RC:: P_hM4D (Ray et al., 2011) muizen om dieren af te leiden, waarbij alle muizen het RC::P_hM4D allel droegen. Verwante dieren die het Cre-allel niet erven, werden gebruikt als controledieren voor de CRE-positieve Nakomelingen. Rosa26 specifieke primers voor de AI9, RC::P_hM4D, en RC::ePe muizen waren 5 ‘- GCACTTGCTCTCCCAAAGTC, 5’-GGGCGTACTGGCATATGAT, en 5 ‘ – CTTAAGCCTGCCCAGAAGA en leveren een 495 bp band (targeted) en 330 BP band (wt). Cre-specifieke primers voor alle rhombomere Cre drivers waren 5 ‘- ATCGCCATCTTCCAGCGCACCATTGCCC en 5 ‘ – GCATTTCTGGGGATTGCTTA en leverde een 550 BP band als positief. Voor LC-MS experimenten, werden c57bl / 6J muizen verkregen van het Center of Comparative Medicine( CCM), Baylor College Of Medicine.
histologie
vier tot acht weken oude DBH-Cre; volwassen muizen van Ai9 werden transcardiaal geperforeerd met 0,1 M fosfaatgebufferde zoutoplossing (PBS) en vervolgens met 4% paraformaldehyde (PFA) in PBS. Hersenen werden ontleed en druppel gefixeerd voor 2 uur in 4% PFA vóór een PBS-spoeling en equilibratie in 20% sucrose in PBS. De hersenen werden toen in secties van 30 µm verdeeld, op dia ‘ s opgezet, en met immunofluorescent antilichamen geëtiketteerd. We bevlekten ‘ s nachts met anti-tyrosine hydroxylase antilichaam om catecholaminerge neuronen te identificeren (1:1.000, Millipore AB152) gevolgd door 2 uur met donkey anti-rabbit CY3 secundair (1:500, Jackson 711-165-152) in 0,1% Triton-X in PBS (PBST) met 5% donkey serum. Beelden werden verzameld op een Zeiss rechtop epifluorescente microscoop.
elektrofysiologie
horizontale hersenplakken met de locus coeruleus (300 µm) werden gesneden met een vibratoom (Leica VT 1000S, Leica Microsystems, Buffalo Grove, IL) van volwassen DBH-Cre; RR2P; RC::Epe muizen (~1 maand oud) in 4°C kunstmatige cerebrospinale vloeistof (ACSF). De plakjes werden ondergedompeld in een kamer geperforeerd met zuurstofrijk ACSF (95% O2 en 5% CO2) met in mM: 124 NaCl, 2,0 KCl, 1,3 MgSO4, 2,5 CaCl2, 1,2 KH2PO4, 25 NaHCO3 en 10 glucose (2-3 ml/min). Opnames van hele cellen werden uitgevoerd bij 30°C met behulp van conventionele patch-clamp technieken en een MultiClamp 700B versterker (Molecular Devices, Union City, CA). GFP-positieve neuronen van locus coeruleus werden visueel geà dentificeerd en later afgebeeld door infrarode differentiële interferentiecontrastvideo op het stadium van een rechte microscoop (Axioskope FS2, Carl Zeiss, Oberkochen, Duitsland). Patchpipetten (weerstanden 4-6 Me) werden gevuld met (in mM): 110 K-gluconaat, 10 KCI, 10 HEPES, 10 Na2-fosfocreatine, 2 Mg3-ATP en 0,2 Na3-GTP; pH werd aangepast tot 7,2 en osmolariteit tot 300 mOsm. Het bedrijfspotentieel was -70 mV. CNO werd bad toegepast.
plethysmografie
plethysmografie op bewuste, ongeremde muizen werd uitgevoerd zoals beschreven bij volwassen dieren van 6 tot 12 weken oud (Ray et al., 2011). Gewone muizen werden onderworpen aan een 5-daagse gewenningsprotocol met elke dag bestaande uit enkele minuten van behandeling, temperatuur genomen door rectale sonde, intraperitoneale zoutoplossing injectie, en 30 minuten in de plethysmografie kamer. Muizen werden vervolgens getest binnen 1 week na de laatste dag van gewenning. Niet-gehabiteerde muizen werden niet blootgesteld aan hantering of de plethysmograafkamer. Alle muizen waren naïef voor CNO en gebruikten slechts één keer.
op de dag van de test werden muizen uit hun kooi genomen, gewogen en werd de rectale temperatuur gemeten. De dieren werden vervolgens in een doorstroom -, temperatuurgecontroleerde (watermantel bij 30°C) plethysmografie-kamer geplaatst en gedurende ten minste 20 minuten in de lucht (21% O2/79% N2) geacclimatiseerd. Na acclimatie en meting onder de kamerlucht werd het kamergas gedurende 20 minuten overgeschakeld op een hypercapnisch mengsel van 5% CO2/21% O2/74% N2. Daarna werd het kamergas gedurende 20 minuten weer op de lucht in de kamer gezet. De muizen werden kort verwijderd voor rectale temperatuurmeting en intra-peritoneale injectie van CNO (National Institute of Mental Health Chemical Synthesis and Drug Supply Program) opgelost in zoutoplossing (1 of 0,1 mg/ml) voor een effectieve concentratie van 10 of 1 mg/kg, respectievelijk. Het dier werd terug naar de kamer gebracht voor nog eens 20 min van lucht in de kamer, 20 min van hypercapnie, en 20 min van lucht in de kamer. Het dier werd vervolgens uit de kamer verwijderd en de rectale temperatuur werd onmiddellijk na afloop van het experiment opgenomen.
vloeistofchromatografie-massaspectrometrie
vierentwintig wildtype muizen, gelijkmatig verdeeld naar geslacht, werden gewogen en behandeld met 10 mg/kg CNO, 1 mg/kg CNO, 0,1 mg/kg CNO of vehiculum. Dertig minuten na de injectie werden bloedmonsters verzameld via een hartpunctie en in bd-Microtainers geplaatst. De monsters werden gecentrifugeerd bij 4°C bij 13.500 tpm in een centrifuge op tafel, en supernatants werden verzameld. Serum en hersenen werden gehouden op -20°C tot extractie.
oplosmiddelen van HPLC-kwaliteit water, acetonitril chloroform en methanol en mierenzuur van massaspectrometrie werden verkregen uit Sigma-Aldrich (St.Louis, MO). Kalibratieoplossing met meerdere kalibranten in een oplossing van acetonitril, trifluorazijnzuur en water werd gekocht bij Agilent Technologies (Santa Cruz, CA). Metabolieten en interne standaarden, waaronder N-acetyl asparaginezuur-d3, tryptofaan-15N2, glutaminezuur-d5, thymine-d4, gibberellinezuur, trans-zeatine, jasmoninezuur, 15N antranilzuur en testosteron-d3 werden gekocht van Sigma-Aldrich (St.Louis, MO). Microtainer R SST TM werd verkregen van Becton Dickinson (Franklin Lakes, NJ).
extractie bestond uit 750 µl ijskoude methanol:water (4: 1) met 20 µl gepikte interne standaarden die werd toegevoegd aan elk hersenmonster (50 mg) en kwaliteitscontroles en vervolgens gedurende 1 min werd gehomogeniseerd. Vervolgens werd 750 µl 100% acetonitril met 20 µl gepikte interne standaarden toegevoegd aan het wasmonster (100 µl) en kwaliteitscontroles en vervolgens gedurende 5 minuten gesoniceerd. Alle monsters werden gecentrifugeerd bij 5000 toeren per minuut gedurende 10 min bij 4°C. De resulterende supernatant werd verzameld en 20 µl werd geïnjecteerd in LC-MS
alle monsters werden geanalyseerd met behulp van 6490 triple quadrupole massaspectrometer (Agilent Technologies, Santa Clara, CA) gekoppeld aan HPLC-systeem (Agilent Technologies, Santa Clara, CA) door middel van multiple reaction monitoring (MRM). Per gedetecteerde metaboliet werden ongeveer 8-11 gegevenspunten verkregen. De gedetecteerde metabolieten waren clozapine, CNO en norclozapine (N-desmethyl clozapine). ESI positieve modus werd gebruikt in methode. De HPLC-kolom was ACQUITYUPLC C18-kolom (100 Å, 1,8 µm en 2,1 mm × 100 mm). Milford, MA, USA) met een debiet van 0,5 ml / min.
gegevensverzameling en-analyse
plethysmografie
drukveranderingen in plethysmografie werden gemeten met behulp van een Validyne dp45 drukverschiltransducer en referentiekamer en CD15 dragerdemodulator en geregistreerd met LabChartPro in real time. Golfvormen werden offline geanalyseerd met behulp van LabChartPro en aangepaste geschreven MATLAB-code om respiratoire snelheid (RR), tidal volume (VT) te bepalen (Ray et al., 2011), minute ventilation (VE) en pattern analysis. Respiratoire golfvormen werden offline verzameld tijdens perioden waarin het dier in rust was, en de metingen waren vrij van bewegingsartefacten. Een minimum van 1 min cumulatieve gegevens samengesteld uit sporen ten minste 10 S Lang van de laatste 10 min van een gegeven experimentele toestand werden geanalyseerd. Het O2-verbruik werd bepaald door de gassamenstelling tussen kalibratie in een lege kamer en levende ademhaling te vergelijken met behulp van een AEI-zuurstofsensor en-analysator. De temperatuur in de kamer werd constant gecontroleerd met behulp van een ThermoWorks MicroThermo 2 en sonde en werd in real time geregistreerd met LabChartPro.
Poincaré metingen en zucht-en apneu frequentie werden bepaald met behulp van 1 min bewegingsvrije sporen van elke ademhalingsaandoening. Zuchten werden gedefinieerd als een adem met een amplitude van minstens twee keer zo groot als de gemiddelde adem. Apneu ‘ s werden gedefinieerd als een interbreath interval (Ibi) ten minste tweemaal zo groot als de gemiddelde IBI. De variatiecoëfficiënt (CV) van de IBI en amplitude werd ook berekend op basis van dezelfde 1-min spoorcompilatie van elke ademhalingsaandoening (standaardfout IBI of amplitude/gemiddelde IBI of amplitude).
statistieken
plethysmografie
resultaten (RR, VT, VE, VO2, VE/VO2, aantal apneu ’s en zuchten, en cv’ s van IBI en amplitude) voor kamerlucht-en hypercapnische gegevens werden vergeleken tussen DBH-Cre; RC::p_hm4d-cohorten en sibling-controles met behulp van een lineair regressiemodel met gemengde effecten met diertype (experimenteel vs.controle) en CNO-toediening (pre vs. post) injectie) als vaste effecten en dierlijke ID als een willekeurig Effect. Temperatuurgegevens werden vergeleken met behulp van een lineair mixed-effect regressiemodel met diertype (experimenteel vs.controle) als een vast effect. Een p < 0,05 werd gebruikt om de statistische significantie aan te geven, en de individuele gegevenspunten, het gemiddelde en de standaardfout van het gemiddelde worden op alle grafieken weergegeven.
vloeistofchromatografie-massaspectrometrie
het verkregen oppervlak onder de piek voor elk monster werd genormaliseerd door de interne controle en vervolgens tot medium voordat statistische analyse werd uitgevoerd. Ongepaarde t-test werd gebruikt om de relatieve abundantie van clozapine en CNO in elk weefsel per concentratiegroep te vergelijken.
ethische verklaring
alle experimenten werden goedgekeurd door de Institutional Animal Care and Use Committee van Baylor College Of Medicine. De proeven voldeden aan de nationale normen voor de verzorging en het gebruik van proefdieren, vastgesteld door de Vereniging voor de beoordeling en accreditatie van Laboratoriumdierverzorging.
Auteursbijdragen
JS, FS-M, MC-M en RR ontwierpen en ontwierpen de experimenten. JS, FS-M en PZ voerden de experimenten uit en droegen bij aan het analyseren van de gegevens. JS, FS-M, VM, MC – M en RR schreven de paper.
financiering
deze studie werd ondersteund door de beurzen R01HL130249 en R01HL130249-S1 van het National Heart, Lung, and Blood Institute; March of Dimes Basil O ‘ Connor Award; en het McNair Medical Institute.
belangenverstrengeling verklaring
de auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Dankbetuigingen
wij danken Drs. Shaila K. Mani en Benjamin Arenkiel voor de nuttige discussies. We danken ook het Baylor College Of Medicine Proteomics Core facility voor de uitvoering van vloeistofchromatografie-massaspectrometrie.Amiel, J., Dubreuil, V., Ramanantsoa, N., Fortin, G., Gallego, J., Brunet, J.-F., et al. (2009). PHOX2B in respiratory control: lessen uit aangeboren centrale hypoventilatie syndroom en zijn muismodellen. Respir. Fysiol. Neurobiol. 168, 125–132. doi: 10.1016 / j.resp.2009.03.005
CrossRef Full Text / Google Scholar
Andrews, N., and File, S. E. (1993). De behandeling van de geschiedenis van ratten wijzigt gedragseffecten van drugs in de verhoogde Plus-doolhof test van angst. Euro. J. Pharmacol. 235, 109–112. doi: 10.1016/0014-2999(93)90827-5
PubMed Abstract / CrossRef Full Text / Google Scholar
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S., and Roth, B. L. (2007). Evoluerend het slot om de sleutel te passen om tot een familie van eiwit-gekoppelde receptoren van G te leiden die krachtig door een inert ligand worden geactiveerd. Proc. Natl. Acad. Sci. USA 104, 5163-5168. doi: 10.1073 / pnas.0700293104
PubMed Abstract / CrossRef Full Text / Google Scholar
Ashby, C. R., and Wang, R. Y. (1996). Farmacologische werking van het atypische antipsychoticum clozapine: een overzicht. Synapse 24, 349-394. doi: 10.1002/(SICI)1098-2396(199612)24:4<349::AID-SYN5>3.0.CO; 2-D
PubMed Abstract / CrossRef Full Text / Google Scholar
Bahia, C. M. C. d. S., and Pereira, J. S. (2015). Obstructieve slaapapneu en neurodegeneratieve ziekten: een bidirectionele relatie. Dement. Neuropsychol. 9, 9–15. doi: 10.1590 / S1980-57642015DN91000003
PubMed Abstract / CrossRef Full Text / Google Scholar
Biancardi, V., Bícego, K. C., Almeida, M. C., en Gargaglioni, L. H. (2008). Locus coeruleus noradrenerge neuronen en CO2 rijden naar de ademhaling. Pflugers Arch. – Euro. J. Physiol. 455, 1119–1128. doi: 10.1007 / s00424-007-0338-8
PubMed Abstract / CrossRef Full Text / Google Scholar
Brust, R. D., Corcoran, A. E., Richerson, G. B., Nattie, E., and Dymecki, S. M. (2014). Functionele en ontwikkelingsidentificatie van een moleculair subtype van hersenen serotonerge neuron gespecialiseerd om de ademhalingsdynamiek te reguleren. Cel Rapport 9, 2152-2165. doi: 10.1016 / j.celrep.2014.11.027
PubMed Abstract / CrossRef Full Text / Google Scholar
Chang, W. H., Lin, S. K., Lane, H. Y., Wei, F. C., Hu, W. H., Lam, Y. W., et al. (1998). Reversibel metabolisme van clozapine en clozapine N-oxide bij schizofrene patiënten. Prog. Neuropsychofarmacol. Biol. Psychiatrie 22, 723-739. doi: 10.1016 / S0278-5846(98)00035-9
PubMed Abstract / CrossRef Full Text / Google Scholar
Chen, Y.-W., Das, M., Oyarzabal, E. A., Cheng, Q., Plummer, N. W., Smith, K. G., et al. (2018). Genetische identificatie van een populatie van noradrenerge neuronen die betrokken zijn bij de verzwakking van stressgerelateerde reacties. Mol. Psychiatrie 1-16. doi: 10.1038 / s41380-018-0245-8
PubMed Abstract / CrossRef Full Text / Google Scholar
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kawasaki, H., Oya, H., Buzza, C., et al. (2015). De ademhaling remde toen de beslagleggingen zich aan amygdala en op Amygdala stimulatie verspreidden. J. Neurosci. 35, 10281–10289. doi: 10.1523 / JNEUROSCI.0888-15. 2015
PubMed Abstract / CrossRef Full Text / Google Scholar
Feinstein, J. S., Buzza, C., Hurlemann, R., Follmer, R. L., Dahdaleh, N. S., Coryell, W. H., et al. (2013). Angst en paniek bij mensen met bilaterale Amygdala schade. Nat. Neurowetenschappen. 16, 270–272. doi: 10.1038 / nn.3323
PubMed Abstract / CrossRef Full Text / Google Scholar
Fitton, A., and Heel, R. C. (1990). Clozapine. Een overzicht van de farmacologische eigenschappen en therapeutisch gebruik bij schizofrenie. Drugs 40, 722-747. doi: 10.2165/00003495-199040050-00007
PubMed Abstract / CrossRef Full Text / Google Scholar
Fleury Curado, T. A., Pho, H., Dergacheva, O., Berger, S., Lee, R., Freire, C., et al. (2018). Het tot zwijgen brengen van hypoglykemische motoneurons leidt tot slaap wanordelijke ademhaling bij magere muizen. Voorkant. Neurol. 9:962. doi: 10.3389 / fneur.2018.00962
CrossRef Full Text / Google Scholar
Gargaglioni, L. H., Hartzler, L. K., and Putnam, R. W. (2010). De locus coeruleus en centrale chemosensitiviteit. Respir. Fysiol. Neurobiol. 173, 264–273. doi: 10.1016 / j.resp.2010.04.024
PubMed Abstract / CrossRef Full Text / Google Scholar
Gomez, J. L., Bonaventura, J., Lesniak, W., Mathews, W. B., Sysa-Shah, P., Rodriguez, L. A., et al. (2017). Chemogenetica onthuld: DREADD bezetting en activering via geconverteerde clozapine. Wetenschap 357, 503-507. doi: 10.1126 / wetenschap.Aan2475
PubMed Abstract / CrossRef Full Text / Google Scholar
Guettier, J. M., Gautam, D., Scarselli, M., Ruiz de Azua, I., Li, J. H., Rosemond, E., et al. (2009). Een chemisch-genetische benadering van studie g eiwitregulatie van bètacelfunctie in vivo. Proc. Natl. Acad. Sci. USA 106, 19197-19202. doi: 10.1073 / pnas.0906593106
CrossRef Full Text / Google Scholar
Hakim, F., Gozal, D., and Kheirandish-Gozal, L. (2012). Sympathische en catecholaminerge veranderingen in slaapapneu met bijzondere nadruk op kinderen. Voorkant. Neurol. 3:7. doi: 10.3389 / fneur.2012.00007
PubMed Abstract / CrossRef Full Text / Google Scholar
Hennessy, M. L., Corcoran, A. E., Brust, R. D., Chang, Y., Nattie, E. E., and Dymecki, S. M. (2017). Activiteit van Tachykinine1-expressie Pet1 raphe neuronen moduleert de respiratoire chemoreflex. J. Neurosci. 37, 1807–1819. doi: 10.1523 / JNEUROSCI.2316-16. 2016
PubMed Abstract / CrossRef Full Text / Google Scholar
Huckstepp, R. T. R., Cardoza, K. P., Henderson, L. E., and Feldman, J. L. (2015). Rol van parafaciale kernen in de controle van de ademhaling bij volwassen ratten. J. Neurosci. 35, 1052–1067. doi: 10.1523 / JNEUROSCI.2953-14. 2015
PubMed Abstract / CrossRef Full Text / Google Scholar
Ilg, A.-K., Enkel, T., Bartsch, D., en Bähner, F. (2018). Gedragseffecten van acute systemische lage dosis clozapine bij wild-type ratten: implicaties voor het gebruik van DREADDs in gedragsneurowetenschap. Voorkant. Behav. Neurowetenschappen. 12:173. doi: 10.3389 / fnbeh.2018.00173
CrossRef Full Text / Google Scholar
Isom, G. E., and Elshowihy, R. M. (1982). Interactie van acute en chronische stress met ademhaling: modificatie door naloxon. Farmacol. Biochem. Behav. 16, 599–603. doi: 10.1016/0091-3057(82)90422-1
PubMed Abstract / CrossRef Full Text / Google Scholar
Jann, M. W., Lam, Y. W., and Chang, W. H. (1994). Snelle vorming van clozapine bij cavia ‘ s en bij de mens na toediening van clozapine-N-oxide. Boog. Int. Pharmacodyn. Ther. 328, 243–250.
PubMed Abstract / Google Scholar
Joober, R. (2010). Clozapine: een duidelijk, slecht begrepen en onderbenut molecuul. J. Neuropsychiatrie. 35, 147–149. doi: 10.1503 / jpn.100055
PubMed Abstract / CrossRef Full Text / Google Scholar
Kinkead, R., Dupenloup, L., Valois, N., and Gulemetova, R. (2001). Stress-geïnduceerde verzwakking van de hypercapnische beademingsreactie bij wakkere ratten. J. Appl. Fysiol. 90, 1729–1735. doi: 10.1152 / jappl.2001.90.5.1729
PubMed Abstract / CrossRef Full Text / Google Scholar
Klein, D. F. (1993). Valse verstikkingsalarmen, spontane paniek en gerelateerde aandoeningen. Een integratieve hypothese. Boog. Gen. Psychiatrie 50, 306-317. doi: 10.1001 / archpsyc.1993.01820160076009
PubMed Abstract / CrossRef Full Text / Google Scholar
Korsak, A., Sheikhbahaei, S., Machhada, A., Gourine, A. V., en Huckstepp, R. T. R. (2018). De rol van parafaciale neuronen in de controle van de ademhaling tijdens inspanning. Sci. Rapport 8: 400. doi: 10.1038/s41598-017-17412-Z
CrossRef Full Text / Google Scholar
Lapin, I. P. (1995). Alleen controles: effect van de behandeling, sham injectie, en Intraperitoneale injectie van zoutoplossing op het gedrag van muizen in een verhoogde plus-doolhof. J. Pharmacol. Toxicol. Methoden 34, 73-77. doi: 10.1016 / 1056-8719 (95)00025-D
PubMed Abstract / CrossRef Full Text / Google Scholar
Lavezzi, A. M., Alfonsi, G., and Matturri, L. (2013). Pathofysiologie van het humane locus coeruleus complex bij foetaal/neonataal plotseling onverklaarbaar overlijden. Neurol. Res. 35, 44-53. doi: 10.1179 / 1743132812Y. 0000000108
PubMed Abstract / CrossRef Full Text / Google Scholar
MacLaren, D. A. A., Browne, R. W., Shaw, J. K., Krishnan Radhakrishnan, S., Khare, P., España, R. A., et al. (2016). Toediening van Clozapine N-oxide veroorzaakt gedragseffecten bij long-evans-ratten: implicaties voor het ontwerpen van DREADD-experimenten. eNeuro 3, 1-14. doi: 10.1523 / ENEURO.0219-16. 2016
PubMed Abstract / CrossRef Full Text / Google Scholar
Madisen, L., Zwingman, T. A., Sunkin, S. M., Oh, S. W., Zariwala, H. A., Gu, H., et al. (2010). Een robuust en high-throughput Cre rapportage en karakterisatie systeem voor de hele muis hersenen. Nat. Neurowetenschappen. 13, 133–140. doi: 10.1038 / nn.2467
PubMed Abstract / CrossRef Full Text / Google Scholar
Mahler, S. V., and Aston-Jones, G. (2018). CNO evil? Overwegingen voor het gebruik van dreadds in gedrags-neurowetenschappen. Neuropsychopharmacology 43, 934-936. doi: 10.1038/npp.2017.299
PubMed Abstract / CrossRef Full Text / Google Scholar
Manvich, D. F., Webster, K. A., Foster, S. L., Farrell, M. S., Ritchie, J. C., Porter, J. H., et al. (2018). De DREADD-agonist clozapine N-oxide (CNO) wordt omgekeerd gemetaboliseerd tot clozapine en veroorzaakt clozapine-achtige interoceptieve stimuluseffecten bij ratten en muizen. Sci. Rapport 8, 3840. doi: 10.1038/s41598-018-22116-Z
CrossRef Full Text / Google Scholar
Misslin, R., Herzog, F., Koch, B., and Ropartz, P. (1982). Effecten van isolatie, behandeling en nieuwigheid op de hypofyse-bijnierreactie bij de muis. Psychoneuroendocrinologie 7, 217-221. doi: 10.1016/0306-4530(82)90015-4
PubMed Abstract / CrossRef Full Text / Google Scholar
Nakamura, A., Zhang, W., Yanagisawa, M., Fukuda, Y., and Kuwaki, T. (2007). Vigilance state-afhankelijke verzwakking van hypercapnic chemoreflex en overdreven slaapapneu in orexin knockout muizen. J. Appl. Fysiol. 102, 241–248. doi: 10.1152 / japplphysiol.00679.2006
PubMed Abstract / CrossRef Full Text / Google Scholar
Obonai, T., Yasuhara, M., Nakamura, T., en Takashima, S. (1998). Catecholamine neuronen verandering in de hersenstam van slachtoffers van wiegendood syndroom. Kindergeneeskunde 101, 285-288. doi: 10.1542 / peds.101.2.285
PubMed Abstract / CrossRef Full Text / Google Scholar
Ozawa, Y., Takashima, S., en Tada, H. (2003). Alpha2-adrenerge receptor subtype veranderingen in de hersenstam bij het wiegendood syndroom. Vroege Zoem. Dev. 75 (Suppl.), S129–S138. doi: 10.1016 / j. pathophys.2004.01.015
CrossRef Full Text / Google Scholar
Padovan-Hernandez, Y., and Knackstedt, L. A. (2018). Dosisafhankelijke vermindering van de door cocaïne geïnduceerde voortbeweging door clozapine-N-oxide bij ratten met een voorgeschiedenis van zelftoediening van cocaïne. Neurowetenschappen. Lett. 674, 132–135. doi: 10.1016 / j.neulet.2018.03.045
PubMed Abstract / CrossRef Full Text / Google Scholar
Ramanantsoa, N., Hirsch, M.-R., Thoby-Brisson, M., Dubreuil, V., Bouvier, J., Ruffault, P.-L., et al. (2011). Ademhaling zonder co (2) chemosensitiviteit bij voorwaardelijke Fox2b-mutanten. J. Neurosci. 31, 12880–12888. doi: 10.1523 / JNEUROSCI.1721-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Raper, J., Morrison, R. D., Daniels, J. S., Howell, L., Bachevalier, J., Wichmann, T., et al. (2017). Metabolism and distribution of clozapine-N-oxide: implications for nonhuman primate chemogenetics. ACS Chem. Neurosci. 8, 1570–1576. doi: 10.1021/acschemneuro.7b00079
PubMed Abstract | CrossRef Full Text | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Soriano, L. P., Nattie, E. E., and Dymecki, S. M. (2013). Egr2-neuronen controleren de volwassen respiratoire respons op hypercapnie. Brain Res. 1511, 115-125. doi: 10.1016 / j. brainres.2012.12.017
PubMed Abstract / CrossRef Full Text / Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Kim, J. C., Richerson, G. B., Nattie, E., et al. (2011). Verminderde controle van de ademhalings-en lichaamstemperatuur na acute serotonerge neuronremming. Science 333, 637-642. doi: 10.1126 / wetenschap.1205295
PubMed Abstract / CrossRef Full Text / Google Scholar
Ryabinin, A. E., Wang, Y. M., and Finn, D. A. (1999). Verschillende niveaus van Fos-immunoreactiviteit na herhaalde hantering en injectiestress bij twee inteelt stammen van muizen. Farmacol. Biochem. Behav. 63, 143–151. doi: 10.1016 / S0091-3057(98)00239-1
PubMed Abstract / CrossRef Full Text / Google Scholar
Snyder, B., Shell, B., Cunningham, J. T., and Cunningham, R. L. (2017). Chronische intermitterende hypoxie induceert oxidatieve stress en ontsteking in hersengebieden geassocieerd met vroege stadium neurodegeneratie. Fysiol. Rapport 5, 1-13. doi: 10.14814 / phy2.13258
PubMed Abstract / CrossRef Full Text / Google Scholar
Sun, J. J., Huang, T.-W., Neul, J. L., and Ray, R. S. (2017). Embryonale hindbrain patterning genen onderscheiden cardio-respiratoire en metabolische homeostatische populaties in de volwassene. Sci. Rep. 7: 9117. doi: 10.1038/s41598-017-08810-4
CrossRef Full Text / Google Scholar
Sun, J. J., and Ray, R. S. (2017). TG (Th-Cre)FI172Gsat (TH-Cre) definieert neuronen die nodig zijn voor volledige hypercapnische en hypoxische reflexen. Biol. Open 6, 1200 1208. doi: 10.1242 / bio.026823
PubMed Abstract / CrossRef Full Text / Google Scholar
Taugher, R. J., Lu, Y., Wang, Y., Kreple, C. J., Ghobbeh, A., Fan, R., et al. (2014). De bedkern van de stria terminalis is kritisch voor angst-gerelateerd gedrag veroorzaakt door CO2 en acidose. J. Neurosci. 34, 10247–10255. doi: 10.1523 / JNEUROSCI.1680-14. 2014
PubMed Abstract / CrossRef Full Text / Google Scholar
Valentino, R. J., and van Bockstaele, E. (2008). Convergente regulatie van locus coeruleus activiteit als een adaptieve reactie op stress. Euro. J. Farmacol. 583, 194–203. doi: 10.1016 / j. ejphar.2007.11.062
CrossRef Full Text / Google Scholar
Verbraecken, J., and McNicholas, W. T. (2013). Ademhalingsmechanica en beademingscontrole bij overlapsyndroom en obesitas hypoventilatie. Respir. Res. 14: 132. doi: 10.1186/1465-9921-14-132
PubMed Abstract / CrossRef Full Text / Google Scholar
Viemari, J.-C. (2008). Noradrenerge modulatie van het neurale netwerk van de luchtwegen. Respir. Fysiol. Neurobiol. 164, 123–130. doi: 10.1016 / j.resp.2008.06.016
PubMed Abstract / CrossRef Full Text / Google Scholar
Winter, A., Ahlbrand, R., Naik, D., en Sah, R. (2017). Differentiële gedragsgevoeligheid voor kooldioxide (CO2) inhalatie bij ratten. Neurowetenschappen 346, 423-433. doi: 10.1016 / j. neurowetenschappen.2017.01.003
PubMed Abstract / CrossRef Full Text / Google Scholar
Ziemann, A. E., Allen, J. E., Dahdaleh, N. S., Drebot, I. I., Coryell, M. W., Wunsch, A. M., et al. (2009). De amygdala is een chemosensor die kooldioxide en acidose detecteert om angstgedrag op te wekken. Cell 139, 1012–1021. doi: 10.1016/j.cell.2009.10.029
PubMed Abstract | CrossRef Full Text | Google Scholar