PUBLICIDAD:
En este artículo discutiremos sobre el sistema digestivo de los vertebrados con la ayuda de diagramas adecuados.
Tracto Digestivo Embrionario:
Archenteron:
El archenteron embrionario se convierte en el revestimiento del tracto digestivo adulto y de todos sus derivados. El mesodermo esplácnico agrega capas de tejido conectivo y músculos lisos alrededor del arquenterón. La invaginación ectodérmica de la cabeza forma el estomodeo que conduce a la cavidad oral, y una invaginación ectodérmica ventral media similar forma el proctodeo, que conduce al intestino posterior.
ANUNCIOS:
El estomago se convierte en la cavidad bucal adulta y da lugar al esmalte de los dientes, la cubierta epitelial de la lengua, glándulas, por ejemplo, mucosas, venenosas y salivales, etc. y la bolsa de Rathke de la glándula pituitaria anterior. El proctodeo forma una pequeña parte terminal de la cloaca en los vertebrados inferiores y el recto en los mamíferos.
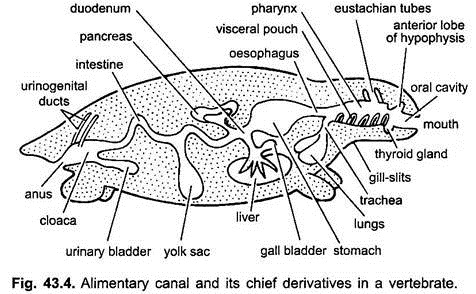
El canal alimentario en embriones desde el estómago hasta la cloaca está unido a la pared dorsal del cuerpo por un doble pliegue de peritoneo, llamado mesenterio dorsal, y a la pared ventral del cuerpo por un mesenterio ventral. En los adultos, el mesenterio dorsal persiste, pero el mesenterio ventral desaparece dejando solo en la región del hígado y la vejiga urinaria.
Tracto Digestivo del adulto:
El tracto digestivo se diferencia para diferentes funciones en las siguientes regiones: boca, cavidad bucal, faringe, esófago, estómago, intestino delgado, intestino grueso y cloaca. Las siguientes excrecencias surgen del tracto digestivo: glándulas orales, bolsa de Rathke, glándula tiroides, hendiduras branquiales, cavidad timpánica, timo y otras glándulas de hendiduras branquiales, tráquea, pulmones, vejiga natatoria, hígado, páncreas, saco vitelino y vejiga urinaria.
ANUNCIOS:
Histología:
La pared del canal alimentario está formada por cuatro capas concéntricas.
Son:
(i) Un peritoneo visceral más externo o capa serosa está hecho de células mesoteliales y capa delgada de tejido conectivo. Falta en el esófago,
PUBLICIDAD:
(ii) Debajo hay una capa muscular formada por fibras musculares lisas dispuestas en fibras musculares circulares internas y longitudinales externas. Entre las dos capas de músculos hay una red de células nerviosas y fibras nerviosas del sistema nervioso autónomo, conocido como plexo mientérico o plexo de Auerbach.
(iii) Debajo de la capa muscular hay una submucosa de tejido conjuntivo con fibras elásticas, grasa, vasos sanguíneos y linfáticos, células nerviosas y glándulas fibrosas,
(iv) La capa más interna es una mucosa compuesta de tres regiones:
(a) Mucosa muscular más estrecha externa de fibras musculares lisas circulares longitudinales externas e internas.
(b) Capa delgada media de lámina propia de tejido conjuntivo, vasos sanguíneos, nervios y nódulos de tejido linfático, y
(c) Una membrana basal que soporta una capa de células epiteliales cilíndricas que a menudo son glandulares y ciliadas.
Boca:
La boca es la abertura que conduce a la cavidad bucal. En lampreas (ciclostomas) es una abertura circular en la base del embudo bucal y permanece permanentemente abierta debido a la falta de mandíbulas, etc. En gnatóstomos es terminal. La boca está limitada por labios que son inamovibles y formados de piel cornificada en peces, anfibios y reptiles. En los mamíferos, estos son carnosos y musculosos.
Cavidad Bucal:
PUBLICIDAD:
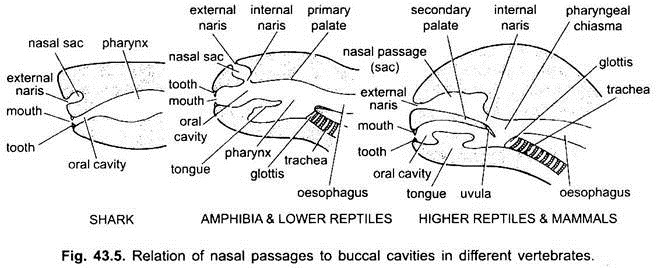
El espacio entre los labios y las mandíbulas es un vestíbulo. Puede estar limitada en el exterior por las mejillas y en el interior por las encías. Las glándulas mucosas de las mejillas se abren en el vestíbulo. La boca se abre en una cavidad bucal, que es un espacio entre la boca y la faringe. El punto exacto donde el ectodermo estomodeal y el endodermo faríngeo se fusionan es variable y no es fácil de discernir.
En los elasmobranquios y la mayoría de los peces óseos, las cavidades nasales no se abren en la cavidad bucal. En los condrichtios y tetrápodos (anfibios y la mayoría de los reptiles), las cavidades nasales se abren en la cavidad bucal por medio de coanas o narinas internas, que se colocan primitivamente en la parte anterior, pero en los cocodrilos, las aves y los mamíferos se vuelven posteriores en la faringe debido a la formación de un paladar secundario, que separa efectivamente el conducto nasal respiratorio de la cavidad bucal o el paso de alimentos.
En las aves, este paladar está hendido debido a que las cavidades nasales y bucales se comunican entre sí. En mamíferos, el paladar secundario se continúa posteriormente como paladar blando membranoso. En los seres humanos, el paladar blando pende de la faringe laríngea en forma de proceso carnoso, llamado úvula.
Derivados de la Cavidad Bucal:
1. Glándulas orales:
Hay dos tipos de glándulas multicelulares integumetarias que se abren en la cavidad bucal. Son glándulas mucosas y glándulas enzimáticas. Los peces y anfibios acuáticos solo tienen glándulas mucosas. Los reptiles tienen glándulas en grupos, como glándulas palatinas, linguales, sublinguales y labiales nombradas de acuerdo con la ubicación, también producen moco.
En las serpientes venenosas, las glándulas labiales superiores se modifican para secretar veneno, mientras que en el monstruo de Gila, las glándulas sublinguales producen veneno. Las aves tienen glándulas sublinguales y una glándula en el ángulo de la boca. Los mamíferos tienen muchas glándulas mucosas pequeñas además de glándulas salivales verdaderas y agrandadas que son enzimáticas. Son glándulas salivales parótidas, sublinguales, submaxilares e infraorbitales, que secretan mucina y ptialina.
2. Lengua:
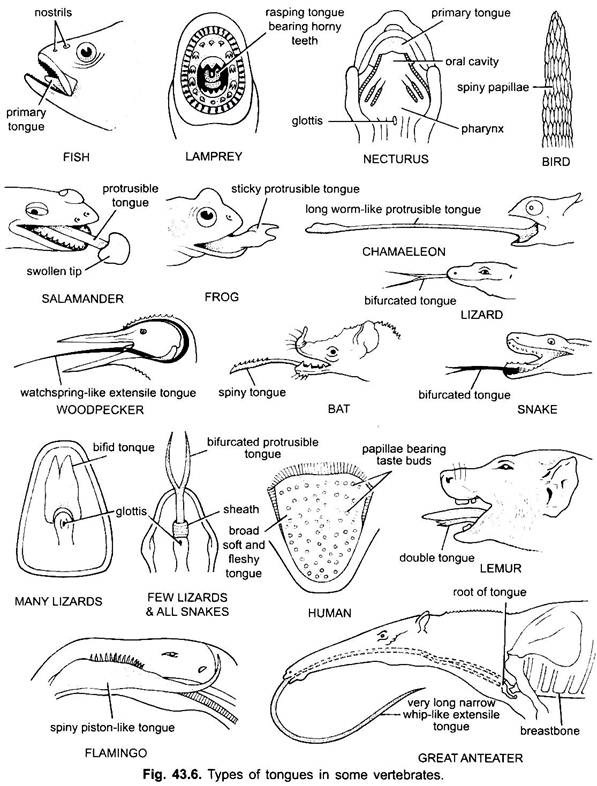
La lengua se encuentra principalmente en todos los vertebrados. La lengua en los vertebrados muestra mucha diversidad y no es homóloga. En los ciclóstomos, hay una lengua musculosa, carnosa y rasposa con dientes córneos para raspar la piel y los músculos de su presa.
Los peces tienen una lengua primaria formada por un pliegue carnoso del suelo bucal. No tiene músculos, pero hay receptores y dientes presentes en la lengua en algunos peces óseos. La lengua está cubierta de membrana mucosa. En algunos anfibios, la lengua es inexistente o inamovible. La mayoría de los anfibios, sin embargo, tienen una lengua protuberante y en algunas ranas y sapos puede plegarse sobre sí misma cuando no está en uso.
Se puede expulsar de la boca mediante una rápida entrada de linfa para capturar los insectos. La lengua de los lagartos y las serpientes a menudo está muy desarrollada. En los camaleones, es muy extensible para capturar insectos. La punta está espesa y pegajosa. La punta bifurcada de la lengua en las serpientes sirve como un medio para transferir estímulos químicos desde el entorno externo a los órganos vomero-nasales emparejados en el techo de la boca.
En tortugas y cocodrilos, la lengua no se puede extender. La lengua amniota tiene músculos voluntarios, recibe el nervio hipogloso y tiene glándulas y papilas gustativas. También desarrolla músculos intrínsecos que mueven la lengua. En las aves, la lengua es delgada y tiene una cubierta córnea. En algunas aves, la lengua es inmóvil, mientras que en algunas aves es larga, protráctil y a menudo se usa para capturar el alimento.
En la mayoría de los mamíferos, excepto las ballenas, la lengua está altamente desarrollada y es capaz de un movimiento considerable, además de la extensión y la retracción, debido a la presencia de una serie de músculos intrínsecos.
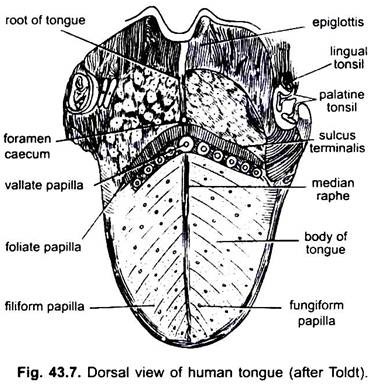
En los mamíferos, la membrana mucosa debajo de la lengua forma un pliegue medio, llamado frenillo, que une la lengua con el suelo de la boca. En los mamíferos, la superficie superior de la lengua tiene cuatro tipos de papilas (filiformes, fungiformes, foliadas y circunvaladas), con papilas gustativas, excepto las papilas filiformes.
3. Dientes:
Los vertebrados tienen dos tipos de dientes unidos a los huesos de la mandíbula: dientes epidérmicos y dientes verdaderos. Los dientes epidérmicos se desarrollan mejor en ciclostomas. Son estructuras córneas duras, cónicas, derivadas del estrato córneo. En las lampreas se encuentran en las paredes del embudo bucal y en la lengua. Las larvas de renacuajos de ranas y sapos tienen dientes epidérmicos serrados en filas en los labios. En los mamíferos, el ornitorrinco adulto tiene dientes epidérmicos.
Dientes verdaderos:
Los dientes no se encuentran en ballenas barbadas y osos hormigueros en mamíferos, agnatos, esturiones, algunos sapos, sirenios,tortugas y aves modernas, etc. En vertebrados inferiores (como peces, anfibios y la mayoría de reptiles) los dientes pueden ser reemplazados continuamente un número indefinido de veces, tales dientes se llaman polifiodontes. Estos dientes son homodontes (tipo similar) y acrodontes. (con los huesos de la mandíbula).
En la mayoría de los mamíferos, los dientes son difiodontes, tecodontes y heterodontes. En algunos mamíferos, estos son monofiodontes que tienen un solo juego de dientes, por ejemplo, lunares, ardilla india. Los dientes tienen una estructura similar a las escamas placoides de los tiburones, formadas por una cavidad pulpar central, alrededor de la cual está presente una capa gruesa pero suave, la dentina, que está cubierta externamente por un esmalte delgado y extremadamente duro. Se supone que se derivan de escamas óseas de ostracodermos y placodermos. Para más detalles, los lectores pueden ver la Dentición en mamíferos.
4. Adenohipófisis:
El lóbulo anterior de la glándula pituitaria se desarrolla como una evaginación dorsal del estomodeo, llamada bolsa de Rathke, que se constriñe para formar los lóbulos anterior y medio de la glándula pituitaria (adenohipófisis). El lóbulo posterior de la hipófisis o neurohipófisis es la evaginación ventral del diencéfalo, llamada infundíbulo. Por lo tanto, es parte nerviosa.
Faringe:
La parte del canal alimentario inmediatamente detrás de la cavidad bucal es una faringe, revestida con endodermo. Es un pasaje que sirve tanto para la digestión y la respiración. Como parte del sistema digestivo, se utiliza como pasaje para los alimentos desde la cavidad bucal hasta el esófago, sus músculos inician la deglución.
En los peces, la faringe es grande y perforada lateralmente para hendiduras branquiales, mientras que en la tetrápoda, es corta y tiene aberturas de fosas nasales. En los embriones, la pared de la faringe emite una serie de evaginaciones que se convierten en espiráculos, hendiduras branquiales, vejigas de aire, pulmones, amígdalas y algunas glándulas endocrinas (por ejemplo, timo, tiroides y paratiroides).
Esófago:
El esófago es corto en la mayoría de los peces y anfibios porque carecen de cuello, pero es más largo en los amnióticos debido a la presencia de cuello. El esófago de los reptiles es más alargado que el de los peces y anfibios. En las aves granívoras y carnívoras, una parte del esófago se agranda en una bolsa en forma de saco llamada cultivo que sirve para almacenar alimentos que se han comido rápidamente.
El cultivo carece esencialmente de glándulas digestivas, aunque en las palomas el cultivo tiene 2 glándulas de cultivo en ambos sexos, en realidad no son glándulas sino estructuras formadoras de células, las células forman «leche de paloma» que se alimenta a los jóvenes. En los mamíferos, el esófago es largo, carece de glándulas y varía en relación con la longitud del cuello.
Pasa a través del diafragma, la porción debajo del diafragma está cubierta con peritoneo visceral que falta en la parte superior. El esófago tiene glándulas mucosas. Su revestimiento forma pliegues longitudinales, o papilas carnosas en forma de dedos (elasmobranquios) o papilas córneas en tortugas marinas.
Histológicamente, el esófago difiere del resto del conducto alimentario en tres hechos:
(i) No tiene peritoneo visceral porque se encuentra fuera del celoma, su capa de cobertura más externa es una túnica adventicia delgada.
(ii) Las fibras musculares en su parte anterior son rayadas, la parte media tiene músculos rayados y no rayados, y la parte posterior solo tiene músculos no rayados. Pero hay excepciones en mamíferos rumiantes, todos los músculos son rayados o voluntarios.
(iii) El revestimiento de la membrana mucosa está hecho de células epiteliales escamosas estratificadas y no de células columnares.
Estómago:
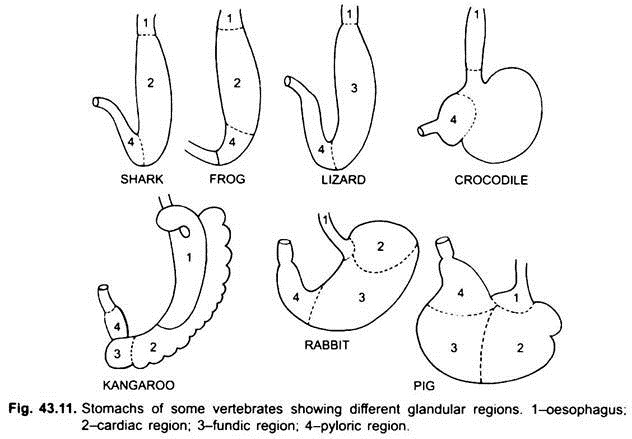
Prácticamente no hay estómago en ciclóstomos, quimeras, peces pulmonares y algunos peces teleósteos primitivos, ya que no tiene glándulas gástricas, pero en la mayoría de los peces y tetrápodos está dilatado para el almacenamiento y la maceración de alimentos sólidos, y la digestión de alimentos porque contiene glándulas gástricas.
La primera parte del estómago, junto al esófago, es la región cardíaca y el extremo inferior cerca del intestino es la región pilórica, que tiene un píloro o válvula pilórica en la que el revestimiento de la membrana mucosa está rodeado por un músculo esfínter grueso que regula la apertura y el cierre del estómago pilórico en el intestino.
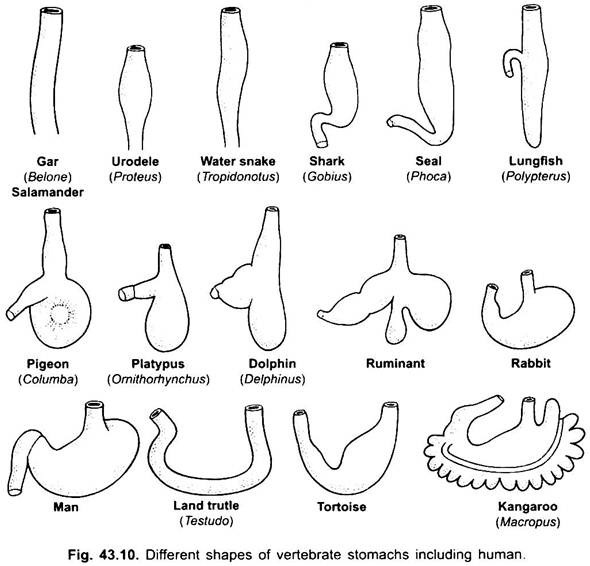
El estómago es recto en ciclostomas, gar, Belone, etc. y con forma de huso en Proteus, Necturus, algunos lagartos y serpientes. En tortugas y tortugas, es un tubo ancho y curvado, y en elasmobranquios el estómago tiene forma de J. En cocodrilos y aves, el estómago tiene dos partes, un proventrículo con glándulas gástricas y una molleja altamente muscular, que representa la región pilórica y tiene un revestimiento duro y cornificado para moler alimentos.
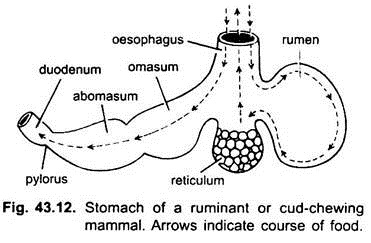
En los mamíferos, el estómago se encuentra transversalmente y puede ser un saco simple o dividido en 3 regiones, a saber, cardíaca, fundica y pilórica, y cada región tiene sus glándulas gástricas. En muchos rumiantes, el estómago tiene cuatro cámaras: el rumen, el retículo, el omaso y el abomaso. Se afirma que las tres primeras cámaras son modificaciones del esófago, y abomaso es el verdadero estómago que representa las partes cardíaca, fundica y pilórica del estómago.
Se ha demostrado embriológicamente que las cuatro cámaras son regiones modificadas del estómago. En los camellos, no hay omaso, el rumen y el retículo tienen células de agua en forma de bolsa que se creía que almacenaban agua, pero probablemente son digestivas.
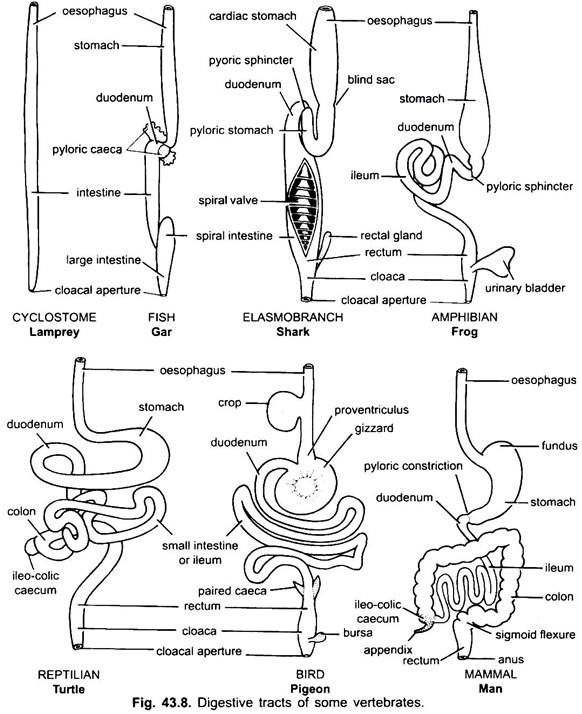
Histológicamente, el estómago tiene las partes típicas del canal alimentario, pero tiene dos peculiaridades, la mucosa muscular está hecha de una capa longitudinal externa y una capa circular interna de músculos. El revestimiento del epitelio es grueso con varios tipos de células glandulares que forman glándulas gástricas de tres tipos llamadas glándulas gástricas cardíacas, fundiculares y pilóricas.
Las glándulas cardíaca y pilórica secretan solamente moco de sus células superficiales. Las glándulas fúndicas (o glándulas cardíacas en algunas) tienen tres tipos de células, las células mucosas del cuello producen moco, las células oxínticas producen ácido clorhídrico, también pueden estar presentes en la región cardíaca, las células zimógenas o las células pépticas secretan pepsina.
En la mayoría de los animales, las células zimógenas también secretan dos proenzimas llamadas propepsina y prorenina, que son convertidas por el ácido clorhídrico en pepsina y renina, respectivamente. Las secreciones de todas las células del estómago forman una mezcla llamada jugo gástrico.
Intestino Delgado:
El intestino delgado es un tubo largo, estrecho y en espiral después del píloro. Es la parte más importante del tracto digestivo porque la digestión y la absorción de los alimentos tienen lugar en él. En los ciclostomas, el intestino es un tubo recto corto con un colgajo longitudinal dispuesto en espiral que se extiende dentro de él.
En elasmobranquios, se divide en porciones pequeñas y grandes, y la porción pequeña tiene una válvula en espiral que aumenta en gran medida la superficie de absorción. Una válvula espiral también está presente en el intestino delgado de algunos peces óseos más primitivos, pero carece de formas superiores en las que el intestino es largo y enrollado.
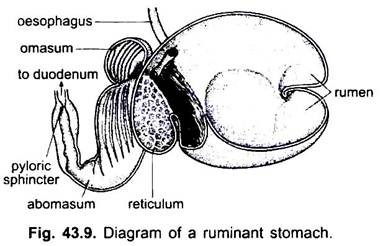
En cecilias, está poco enrollado y no se diferencia en un tracto pequeño y grande. En ranas y sapos es relativamente largo y enrollado. En los reptiles, es más enrollada que en los anfibios. Por primera vez en los vertebrados surge un ciego o divertículo ciego en la unión de los intestinos delgado y grueso.
Sin embargo, esto no es permanente en todos los reptiles. En las aves, el intestino delgado está en espiral o en bucle y uno o dos cólicos ciegos también están presentes en la unión de los intestinos delgado y grueso. En la mayoría de los mamíferos también el intestino delgado es proporcionalmente largo y enrollado. Su longitud, sin embargo, está correlacionada con los hábitos alimenticios. En los herbívoros es relativamente más largo en comparación con los insectívoros y carnívoros.
Hay una bolsa ciega o ciego en la unión del colon y el intestino delgado que generalmente es pequeña en las especies carnívoras y bastante larga en muchos herbívoros. La primera parte del intestino delgado es el duodeno, que es corto a partir del píloro y termina más allá de la entrada de los conductos pancreáticos y hepáticos.
Tiene muchas vellosidades plegadas y contiene glándulas ramificadas de Brunner en la submucosa que secretan moco, algo de líquido acuoso alcalino y un poco de enzima. El duodeno también produce dos hormonas llamadas secretina y colecistoquinina que estimulan el páncreas y la vesícula biliar para liberar sus jugos. Los conductos de la vesícula biliar y el páncreas se abren hacia el duodeno.
Detrás del duodeno hay un ieon, que solo en mamíferos se diferencia en un yeyuno anterior más pequeño y un ieon posterior más largo. Un gran número de pequeñas glándulas digestivas están presentes en el intestino delgado. Son glándulas tubulares o criptas de Lieberkuhn que se encuentran a lo largo de toda la longitud, secretan moco y un succus entericus que tiene varias enzimas.
El revestimiento del intestino delgado se pliega para formar pequeñas vellosidades, que aumentan el área de superficie para la secreción y absorción. Las vellosidades están densamente cubiertas por diminutas proyecciones en forma de dedos, llamadas microvellosidades que ayudan a la absorción en las vellosidades. En los mamíferos se encuentran nódulos de tejido linfoide llamados parches de Peyer en elleumeon.
Intestino grueso:
intestino grueso tiene un diámetro mayor que el del intestino delgado. Generalmente es corto en peces, anfibios, reptiles y aves, pero en mamíferos es largo. En las formas inferiores, el intestino grueso forma un recto, pero en la tetrápoda tiene un colon y un recto terminal. En la mayoría de los peces y anfibios, la parte terminal del recto conduce a una cloaca formada por el proctodeo.
El recto, los conductos excretores y los conductos genitales se abren hacia la cloaca, y se abre hacia el exterior por una abertura cloacal. Pero en muchos peces óseos y en todos los mamíferos (excepto los monotremas), el recto y los conductos urogenitales tienen aberturas separadas hacia el exterior; la abertura de la primera es un ano.
El recto de los mamíferos no es homólogo con el recto de los vertebrados, ya que en los mamíferos se deriva de la partición de la cloaca embrionaria. En la mayoría de los embriones de vertebrados hay un intestino postanal como una extensión del intestino en la cola, pero desaparece más tarde.
En los elasmobranquios, el intestino grueso tiene un par de glándulas rectales que secretan moco y cloruro de sodio. En los amnióticos hay una válvula ileocólica en la unión de los intestinos delgado y grueso, que está ausente en los peces. Evita que las bacterias entren en el íleon de colon.
En los amnióticos de esta unión surge un ciego ileocólico que es dos en las aves. Que contiene las bacterias para digerir la celulosa. Es muy largo, en mamíferos herbívoros (conejo, caballo, vaca, etc.). En primates, el ciego es pequeño y tiene un apéndice vermiforme vestigial.
Glándulas Digestivas:
1. Hígado:
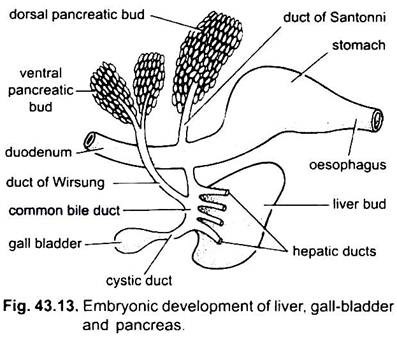
El hígado surge como una consecuencia simple o doble de la pared ventral del arcenterón embrionario. Esta consecuencia forma un divertículo hepático hueco, que pronto se diferencia en una parte anterior, que prolifera para convertirse en el hígado y sus conductos biliares, y una parte posterior, que da lugar a la vesícula biliar y el conducto cístico. Los conductos biliares se unen para formar un conducto hepático que se une con el conducto cístico para formar un conducto biliar común o conducto colédoco. La región del arcenterón de la que surge el hígado se convierte en el duodeno.
El hígado es la glándula lobulada más grande del cuerpo, suspendida por una doble capa de peritoneo del tabique transversal o su representante.
Una vesícula biliar es para el almacenamiento de la bilis secretada por las células hepáticas, se encuentra en el hígado y drena hacia el duodeno a través del conducto biliar común formado por la unión del conducto cístico y el conducto hepático. La vesícula biliar no es indispensable y falta en muchas aves y mamíferos.
El hígado está presente en todos los vertebrados. En los ciclostomas, es pequeño, de un solo lóbulo (lampreas) y de dos lóbulos en los peces bruja. Es bilobulado en elasmobranquios, dos o tres lobulados en peces óseos, anfibios, reptiles y aves y muchos lobulados en mamíferos. El hígado es largo, estrecho y cilíndrico en los peces, urodelos y serpientes.
es corto, ancho y aplanado en aves y mamíferos. La vesícula biliar y el conducto biliar están presentes en los ciclostomas larvarios, pero están ausentes en el adulto. Los peces, anfibios y reptiles generalmente tienen vesícula biliar, pero es deficiente en muchas aves. La mayoría de los mamíferos tienen vesícula biliar, pero está ausente en Cetáceos y Ungulados.
El hígado segrega una bilis acuosa y alcalina, pero no tiene enzimas. Neutraliza la acidez de los alimentos que entran en el duodeno. Ayuda en la digestión de las grasas.
2. Páncreas:
El páncreas se forma a partir del endodermo del arcenteron embrionario. Un único divertículo dorsal del duodeno embrionario y una o dos excrecencias ventrales del hígado forman divertículos pancreáticos. Las partes proximales de los divertículos forman conductos pancreáticos, pero estos conductos se reducen o fusionan de modo que solo uno o dos conductos pancreáticos permanecen en el adulto, se abren al duodeno por separado o después de unirse con el conducto biliar común.
Las partes distales de los divertículos se someten a brotes para formar la masa principal de células pancreáticas a las que se agregan derivados mesodérmicos. Por lo tanto, se forma una sola glándula que tiene varios lóbulos formando un páncreas difuso o compacto.
El páncreas es una glándula exocrina y endocrina, unidas por delicadas hebras de tejido conectivo. La parte exocrina secreta enzimas digestivas que se vierten en el duodeno a través de los conductos pancreáticos. Mientras que la parte endocrina secreta hormonas como la insulina y el glucagón.
El páncreas está presente en todos los vertebrados. En lampreas, algunos peces óseos, peces pulmonares y tetrápodos inferiores, es un órgano difuso incrustado en el hígado, los mesenterios y la pared intestinal. Las hagfish tienen un páncreas pequeño. Los elasmobranquios tienen un páncreas bilobulado bien definido. En la tetrápoda superior, generalmente es una glándula compacta. Uno o dos conductos pancreáticos se abren hacia el duodeno.