PUBLICIDAD:
En el sistema circulatorio de los vertebrados hay dos sistemas de tubos elaboradamente ramificados, que se ramifican por todo el cuerpo y transportan líquidos a los tejidos. Son un sistema vascular sanguíneo y un sistema linfático. Cuando se usa el término sistema circulatorio, se refiere solo al sistema vascular sanguíneo.
El sistema vascular sanguíneo es un sistema cerrado en los vertebrados; tiene un corazón contráctil y tubos continuos llamados vasos. El sistema linfático es un sistema abierto con espacios linfáticos. Los vasos sanguíneos que transportan la sangre desde el corazón son arterias, que se dividen en arteriolas más delgadas, ramificándose en capilares extremadamente delgados y pequeños.
La pared de un capilar está hecha de una sola capa de células endoteliales borlas. Cada vaso sanguíneo, incluido el corazón, tiene un revestimiento de células endoteliales o endotelio. La sangre no entra en contacto directo con las células tisulares.
ANUNCIOS:
Las sustancias pasan desde y hacia los capilares a través del líquido tisular contenido en los espacios tisulares entre las células. El intercambio de sustancias entre la sangre y las células tisulares del cuerpo tiene lugar a través de las paredes capilares. Esto es provocado por la pinocitosis en las células endoteliales de los capilares.
Bajo un microscopio electrónico, las células endoteliales muestran muchas vesículas pequeñas que son invaginaciones de las membranas plasmáticas. Estas vesículas se mueven desde un lado de la célula y liberan su contenido en el lado opuesto.
por Lo tanto, la sangre da oxígeno, material de nutrientes y hormonas a las células, el metabolismo, y las células dar agua, dióxido de carbono y desechos nitrogenados en la sangre para transportar a los órganos excretores para su rápida eliminación.
Los capilares forman una red en todos los tejidos del cuerpo, excepto el cartílago y el epitelio. De los capilares, la sangre pasa a vénulas delgadas que se combinan para formar venas que transportan la sangre hacia el corazón. Pero algunas venas (venas porta, venas renales y venas hepáticas) tienen capilares que son iguales a los de las arterias. Pero toda la sangre no pasa a través de los capilares hacia las vénulas.
ANUNCIOS:
También hay algunos canales a través entre las arteriolas y las vénulas que se encuentran en algunos órganos, como la piel. También hay anastomosis arteriovenosas entre las arteriolas y las vénulas que se encuentran en los dedos. La función de tales conexiones no se conoce claramente, aunque se afirma que regulan la presión arterial y la circulación.
En algunos lugares, el intercambio de material entre los tejidos y la sangre tiene lugar a través de espacios de paredes delgadas o sinusoides. En ciertos órganos, un vaso sanguíneo puede formar una red en espiral de vasos sanguíneos diminutos llamados rete mirabile (riñones, vejiga de aire).
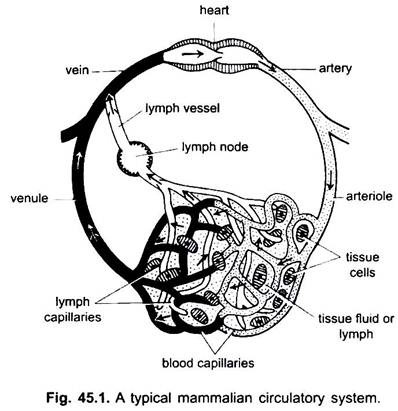
Partes del Sistema Circulatorio:
PUBLICIDAD:
En los cordados, el sistema circulatorio es de tipo cerrado. Este tipo también se encuentra en los anélidos (invertebrados). Otro tipo es de tipo abierto en el que no se encuentran capilares. Se encuentra en moluscos y artrópodos. Aquí la sangre fluye a través de las arterias hacia varios órganos, que pasan a través de los espacios sanguíneos o senos paranasales y luego de nuevo a los vasos (venas) hasta el corazón.
El sistema circulatorio incluye el corazón, las arterias, las venas, los capilares y la sangre. El corazón es un vaso sanguíneo modificado con paredes musculares, que se contrae periódicamente para bombear la sangre a las diversas partes del cuerpo a través de vasos definidos.
Las arterias y sus ramas forman un sistema arterial que transporta la sangre desde el corazón. Las venas y sus afluentes constituyen un sistema venoso que toma la sangre de los capilares de las arterias o arteriolas y la lleva al corazón.
Sistema portal:
En el sistema portal, la sangre no se devuelve directamente al corazón, pero hay un órgano que se interpone (hígado o riñón) en el curso de la sangre que regresa. La vena que trae sangre comienza en los capilares y termina en los capilares, la vena en cuestión actúa como vaso aferente y eferente, los vasos aferentes terminan en los capilares al igual que las arterias, luego la sangre se recolecta en venas sistémicas.
Todos los vertebrados tienen un sistema portal hepático en el que la sangre pasa a dos grupos de capilares en el hígado. Los vertebrados inferiores y los embriones de los vertebrados superiores tienen un sistema portal renal también en el que la sangre pasa a través de dos grupos de capilares en el riñón antes de llegar al corazón. Los capilares de la glándula pituitaria forman un sistema portal pituitario, que es un sistema pequeño pero importante.
Sistema linfático:
Se encuentra en cordados, excepto en ciclostomas y peces cartilaginosos. Incluye linfa y vasos linfáticos. La linfa es un líquido tisular que se encuentra entre las células del cuerpo. Es plasma sanguíneo menos glóbulos rojos y algunas proteínas.
Capilares linfáticos que forman una red de vasos delgados con terminación ciega, que recogen la linfa. Los vasos linfáticos son vasos de paredes delgadas formados por la unión de capilares sanguíneos. Estos se vacían en venas. Los ganglios linfáticos se encuentran en los vasos linfáticos de los mamíferos. Estos forman linfocitos de sangre utilizados para la defensa del cuerpo contra las enfermedades.
Evolución del Corazón en vertebrados:
PUBLICIDAD:
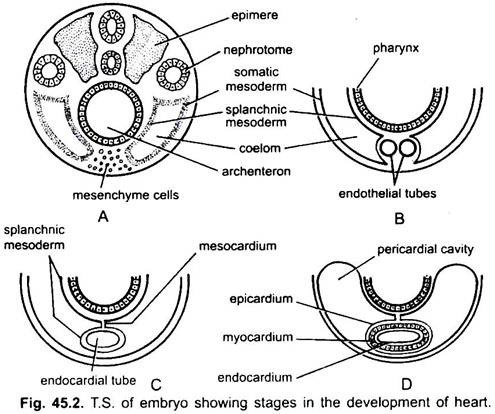
El corazón es un órgano no emparejado, pero su origen es bilateral. En un embrión, el mesénquima forma un grupo de células endocárdicas debajo de la faringe. Estas células se arreglan para formar un par de tubos endoteliales delgados. Los dos tubos endoteliales pronto se fusionan para formar un único tubo endocárdico que se encuentra longitudinalmente debajo de la faringe.
El mesodermo esplácnico situado debajo del endodermo se pliega longitudinalmente alrededor del tubo endocárdico. Este tubo de dos capas formará el corazón en el que el mesodermo esplácnico se espesa para formar un miocardio o pared muscular del corazón y un epicardio delgado externo o pericardio visceral. La sonda endocárdica se convierte en el revestimiento del corazón conocido como endocardio.
Los pliegues del mesodermo esplácnico se unen arriba para formar un mesocardio dorsal que suspende el corazón en el celoma. Pronto se forma un tabique transversal detrás del corazón que divide el celoma en dos cámaras, una cavidad pericárdica anterior que encierra el corazón y una cavidad abdominal posterior.
El corazón es un tubo recto, pero aumenta de longitud y se vuelve en forma de S porque sus extremos son fijos. La aparición de válvulas, constricción, particiones en el corazón y engrosamientos diferenciales de sus paredes forman tres o cuatro cámaras en el corazón.
1. Corazón de una sola cámara:
En el Anfioxo (cordado primitivo), no se encuentra un corazón verdadero. Una parte de la aorta ventral debajo de la faringe es muscular y contráctil y actúa como corazón.
2. Corazón de Dos Cámaras:
En los ciclostomas, hay cuatro cámaras dispuestas en un orden lineal: un venoso sinusal de paredes delgadas, una aurícula ligeramente muscular( pabellón auricular), un ventrículo muscular y un cono arterioso muscular o bulbo cordis. Se encuentra en la cavidad corporal en la que también están presentes otros órganos viscerales.
De las cuatro cámaras, solo la aurícula y el ventrículo corresponden a las cuatro cámaras (aurículas y ventrículos pareados) de los vertebrados superiores. En la evolución del corazón han tenido lugar muchos cambios.
Elasmobranquios:
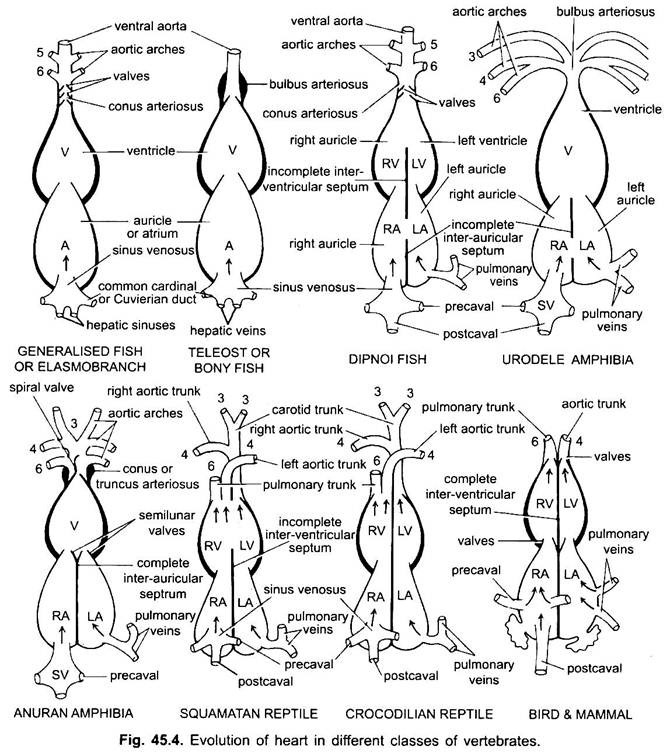
Excepto Dipnoi, el sistema circulatorio en los peces desde los ciclóstomos hasta los teleósteos, solo la sangre no oxigenada va al corazón, desde allí se bombea a las branquias, se airea y luego se distribuye al cuerpo. El corazón del cazón cartilaginoso es un tubo en forma de S musculoso y doblado dorsoventralmente con cuatro compartimentos en una serie lineal.
Son el venoso sinusal y la aurícula para recibir sangre venosa, y un ventrículo y un cono arterioso para bombear esta sangre. El corazón es un corazón venoso branquial. El seno venoso y el cono arterioso son cámaras accesorias. La aurícula y el ventrículo son cámaras verdaderas, por lo tanto, es un corazón de 2 cámaras.
El seno venoso se abre hacia delante en la aurícula a través del río sinú-auricular apertura custodiado por un par de válvulas. La aurícula se encuentra dorsal al ventrículo y se abre ventralmente hacia el ventrículo a través de una abertura auriculoventricular protegida por un par de válvulas. El ventrículo muscular de paredes gruesas se abre en un cono arterioso estrecho que contiene válvulas en dos series.
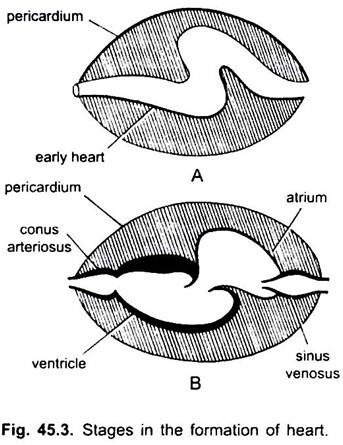
El corazón está encerrado dentro de la cavidad pericárdica separada de la cavidad corporal por un tabique transversal. El cono perfora el pericardio y se vuelve continuo con la aorta ventral. La cavidad pericárdica se comunica con la cavidad corporal a través de dos perforaciones en el tabique transversal.
Teleosts:
Su corazón se asemeja al de los clasmobranquios. En teleósteos, el cono se reduce y tiene un solo par de válvulas. La parte proximal de la aorta ventral cerca del cono se agranda mucho y tiene paredes gruesas, lo que se denomina arterioso bulboso. Es elástico y se dilata en el momento de la contracción ventricular. El corazón es, por lo tanto, de 2 cámaras con una sola circulación de sangre.
3. Corazón de tres cámaras:
Dipnoi:
En diphoans, un tabique divide la aurícula en una cámara derecha e izquierda. Esto se correlaciona con el uso de la vejiga natatoria como órgano de respiración y representa el primer paso hacia el desarrollo del sistema circulatorio de doble tipo, en el que la sangre oxigenada y no oxigenada ingresa al corazón y se mantiene separada.
La sangre de la aurícula derecha del pez pulmón pasa al ventrículo derecho y luego es bombeada a la vejiga de gas primitiva similar a un pulmón por las arterias pulmonares que se ramifican desde el sexto par de arcos aórticos. La sangre oxigenada regresa a la aurícula izquierda a través de venas pulmonares como los anfibios.
Anfibios:
En anfibios, la aurícula dorsal se desplaza anterior al ventrículo. El seno venoso se abre en la aurícula derecha dorsalmente y no posteriormente. La aurícula está completamente dividida en cámaras derecha e izquierda y no tiene foramen oval en el tabique interauricular, que permanece abierto en dipnoans.
Se desarrollan bolsas profundas en la cavidad ventricular. El cono arterioso se divide en vasos sistémicos y pulmonares mediante una válvula espiral. En las salamandras sin pulmón, el tabique interauricular está incompleto y las venas pulmonares están ausentes.
Reptilia:
En los reptiles, el corazón está más avanzada. La aurícula siempre está completamente separada en una cámara derecha e izquierda, y en muchas formas el seno venoso se incorpora a la pared de la aurícula derecha. El ventrículo también está dividido en parte por un tabique en la mayoría de los reptiles, y en los caimanes y cocodrilos es completamente de dos cámaras.
Esto significa que la sangre oxigenada procedente de los pulmones hacia el lado izquierdo del corazón se separa esencialmente de la sangre no oxigenada del cuerpo hacia el lado derecho. Por lo tanto, en los crocodilianos, los dos tipos de sangre están completamente separados, y casi completos en otros reptiles, pero algunas mezclas ocurren en otras partes del sistema circulatorio.
El cono arterioso embrionario se divide en tres vasos en lugar de dos:
(i) Arco pulmonar que lleva sangre a los pulmones desde el lado derecho del ventrículo.
(ii) Aorta sistémica derecha que transporta sangre desde el lado izquierdo del ventrículo hasta el cuerpo a través del cuarto arco aórtico derecho.
(iii) El sistema izquierdo viene del ventrículo derecho al cuarto arco aórtico izquierdo.
En el punto de contacto con la aorta sistémica desde el ventrículo izquierdo, incluso en crocodilianos, está presente una abertura entre los dos, llamada foramen de Panizzae, donde puede haber alguna mezcla de los dos tipos de sangre. Por lo tanto, el corazón de reptil representa el corazón de transición contra el corazón de anfibio: 2 aurículas completas y 2 ventrículos incompletos con una pequeña mezcla de sangre en el sistema derecho e izquierdo.
4. Corazón de cuatro cámaras:
Aves y Mamíferos:
En las aves, el ventrículo se divide completamente en dos, de modo que el corazón tiene cuatro cámaras (2 aurículas y 2 ventrículos). Hay una separación completa de la sangre venosa y arterial. La aorta sistémica sale del ventrículo izquierdo y transporta sangre a la cabeza y al cuerpo. Mientras que la arteria pulmonar sale del ventrículo derecho y lleva sangre a los pulmones para la oxigenación.
Por lo tanto, hay doble circulación en la que no hay mezcla de sangre en ningún lugar. El seno venoso está completamente incorporado en el pabellón auricular derecho, que recibe dos precavales y un postcaval. La aurícula izquierda recibe sangre oxigenada a través de las venas pulmonares, el cono arterioso está ausente, la aorta pulmonar surge del ventrículo derecho y la aorta sistémica única surge del ventrículo izquierdo, y ambos tienen válvulas en sus bases.
Modificaciones de los Arcos Aórticos en vertebrados:
Arterias embrionarias:
Cuando el corazón se está formando en un embrión vertebrado, un vaso sanguíneo llamado aorta ventral aparece en la parte media ventral por debajo de la faringe, que pronto se conecta al cono arterioso. La aorta ventral que surge del corazón corre hacia adelante por debajo de la faringe y se divide anteriormente en un par de arterias carótidas externas en la cabeza.
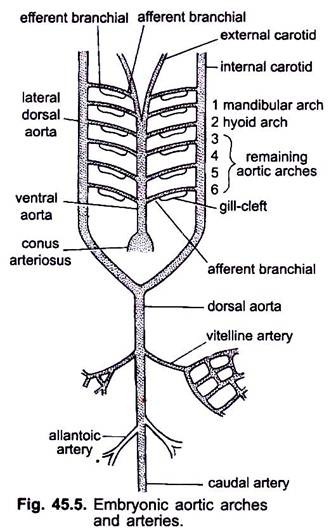
La aorta ventral emite 6 pares de arcos aórticos laterales a equidistancia que atraviesan los arcos viscerales. Cada arco aórtico tiene una arteria branquial aferente ventral que transporta sangre venosa a la branquia y una arteria branquial eferente dorsal que toma sangre oxigenada de la branquia. Las arterias branquiales eferentes de cada lado se unen dorsalmente con la aorta dorsal lateral o aorta radix que entra en la cabeza como arteria carótida interna.
El primer arco aórtico es un arco aórtico mandibular, el segundo es un arco aórtico hioides, los restantes se denominan arcos aórticos tercero, cuarto, quinto y sexto. La aorta dossal lateral se fusiona detrás de la faringe para formar una aorta dorsal que se continúa en la parte media dorsal dentro de la cola como una arteria caudal.
De la aorta dorsal surgen arterias emparejadas y no emparejadas que suministran varios órganos del cuerpo. En un embrión con un saco vitelino, un par de arterias vitelinas surgen de la aorta dorsal y abastecen el saco vitelino. En embriones de amnióticos, un par de arterias umbilicales o alantoicas surge de la aorta dorsal que suministra sangre a los alantois.
En un adulto, las arterias vitelinas se fusionan para formar la arteria mesentérica principal, la mayor parte de las arterias alantoicas se pierde, pero sus restos forman arterias hipogástricas o ilíacas internas.
Arcos aórticos en vertebrados:
En varios vertebrados adultos, el sistema arterial parece ser diferente, pero están construidos sobre el mismo plan fundamental básico. La diferencia se debe a la creciente complejidad del corazón debido a un cambio en la respiración de las branquias a los pulmones. Hay una reducción progresiva en el número de arcos aórticos en la serie de vertebrados.
Cyclostomata:
En Petromyzon, hay 7 pares de arcos aórticos. En otros ciclostomas estos varían de 6 pares en Mixina y 15 pares en Eptatretus.
Piscis:
Aunque se considera que seis han sido el número básico de arcos aórticos para peces. Este número se reduce a cinco incluso en tiburones y rayas con la pérdida de la primera pareja, el arco aórtico mandibular o está representado por una arteria pseudobranquial eferente. En la mayoría de los peces óseos, tanto el arco aórtico mandibular (i) como el hioides (ii) desaparecen o están muy reducidos.
En Polypterus y Dipnoi( peces pulmón), las branquias no están bien desarrolladas. Por lo tanto, la arteria pulmonar surge de la parte eferente del 6º arco a cada lado que suministra sangre a la vejiga de aire o al pulmón. En elasmobranquios y Dipnoi, cada arco tiene una arteria branquial aferente y dos arterias branquiales eferentes (formadas por división) en cada branquias. En los peces óseos, cada branquias tiene una arteria aferente y una eferente.
En la tetrápoda, los arcos aórticos no se rompen en partes aferentes y eferentes porque las branquias internas verdaderas están ausentes. En toda tetrapoda desaparecen el primer y el segundo arco.
Anfibios:
Aquí los arcos aórticos muestran modificaciones debido a la pérdida de branquias y la apariencia de los pulmones. En urodelos hay branquias externas presente como órganos respiratorios, además de los pulmones. Los arcos aórticos III, IV, V y VI están presentes, aunque el quinto par está muy reducido en Sirena, Anfiuma y Necturus. Los arcos aórticos no se rompen en las branquias externas en porciones aferentes y eferentes, porque las ramas que surgen de los arcos aórticos IV, V y VI forman capilares en las branquias externas.
Las aórtas dorsales laterales entre los arcos aórticos III y IV persisten como una conexión vascular, el conducto carótico. El arco aórtico VI forma el arco pulmo-cutáneo o arteria a cada lado que lleva sangre al pulmón y la piel. También retiene una conexión con la aorta dorsal lateral conocida como conducto arterioso (conducto de los Botalos).
En las larvas de anuros (ranas renacuajos), la disposición de los arcos aórticos son como un adulto urodelos debido a la presencia de branquias. En la metamorfosis, con la pérdida de branquias, los arcos aórticos I, II y V desaparecen por completo, solo están presentes los arcos aórticos IIIrd, IV y Vlth. La aorta dorsal lateral entre el tercer y cuarto arco aórtico (conducto carótico) también desaparece. Por lo tanto, el tercer arco aórtico junto con una parte de la aorta ventral se convierte en el arco carotídeo que transporta sangre oxigenada a la región de la cabeza.
El cuarto arco aórtico a lo largo de su aorta dorsal lateral forma el arco sistémico. El sexto arco aórtico se convierte en el arco pulmocutáneo que suministra sangre venosa a los pulmones y la piel. El conducto arterioso desaparece durante la metamorfosis. Por lo tanto, los anuros adultos solo tienen arcos aórticos III, IV y VI. Estos también son retenidos por los amnióticos.
Reptilia:
En los reptiles, las branquias son completamente reemplazadas por los pulmones. Solo están presentes los arcos aórticos III, IV y VI. Con la separación parcial del ventrículo en dos partes, la porción distal del cono arterioso y toda la aorta ventral se dividen en tres vasos, es decir, dos aórticos o sistémicos y uno pulmonar. El arco sistémico derecho (IV) surge del ventrículo izquierdo llevando sangre oxigenada al arco carotídeo (III). Las aórticas sistémicas izquierdas (IV) y pulmonares (VI) tienen su origen en el ventrículo derecho. El sistema izquierdo transporta sangre desoxigenada o mixta al cuerpo a través de la aorta dorsal.
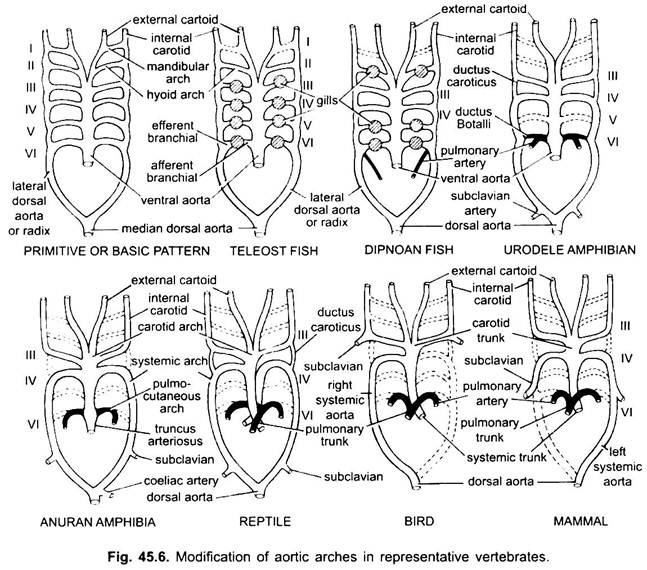
Mientras la arteria pulmonar lleva sangre desoxigenada a los pulmones. El ductus caroticus desaparece, pero persiste en serpientes y algunos lagartos (Uromastix). El conducto arterioso desaparece en la mayoría de los reptiles, aunque persiste en una forma reducida en Sphenodon y algunas tortugas. Debido a la mezcla de sangre, los reptiles son animales de sangre fría como peces y anfibios.
Aves:
En las aves, están presentes los arcos aórticos III, IV y VI. Siguen el patrón general de los reptiles con algunas diferencias. Con la división completa del ventrículo en dos partes, el cono arterioso y la aorta ventral se han dividido para formar dos vasos, la aorta sistémica que surge del ventrículo izquierdo y la aorta pulmonar del ventrículo derecho.
El tercer arco aórtico con restos de aorta lateral y ventral forma las carótidas que surgen de la aorta sistémica. El cuarto arco aórtico forma la aorta sistémica solo en el lado derecho. Se une con la aorta lateral de su propio lado y forma la aorta dorsal. Parte del cuarto arco aórtico del lado izquierdo forma la arteria subclavia izquierda, el resto junto con su aorta dorsal lateral desaparece. El sexto arco aórtico forma la aorta pulmonar. El conducto carótico y el conducto arterioso desaparecen.
Mammalia:
En mamíferos también persisten los arcos aóricos III, IV y VI. El ventrículo se divide completamente en dos partes.
El cono arterioso y la aorta ventral se dividieron para formar dos vasos:
(i) Una aorta sistémica que surge del ventrículo izquierdo, y
(ii) Una aorta pulmonar del ventrículo derecho. El tercer arco aórtico con restos de aorta lateral y ventral forma el arco carotídeo.
El cuarto arco aórtico forma la aorta sistémica solo en el lado izquierdo, mientras que en el lado derecho su porción proximal forma una arteria subclavia innominada y derecha, el resto junto con su aorta dorsal lateral desaparece. El sexto arco aórtico forma la aorta pulmonar. El conducto arterioso degenera, pero persiste en algunos hasta la eclosión o el nacimiento en una forma reducida en el lado izquierdo como un ligamento arterioso delgado.
Sistema venoso:
Venas Embrionarias:
En todos los embriones vertebrados, el sistema venoso es simple y similar. Las venas están en su mayoría emparejadas y dispuestas simétricamente. En embriones sin saco vitelino, se forma una vena sub-intestinal en el mesodermo esplácnico debajo del intestino.
Se repite alrededor del ano y se continúa posteriormente como:
(i) Una vena caudal en la cola. En todos los embriones que tienen un saco vitelino (ya sea que lo contengan o no), un par de venas vitelinas surgen del saco vitelino y se unen a la parte posterior del corazón en desarrollo, que se convierte en el seno venoso. De hecho, la fusión de estas venas vitelinas es responsable de la formación del corazón en peces óseos, reptiles y aves. Cada vena vitelina en su extremo posterior se une a la vena subintestinal formada de la misma manera que en los embriones que no tienen saco vitelino.
(ii) Un par de venas subcardinales surgen entre los riñones y se unen a la vena caudal.
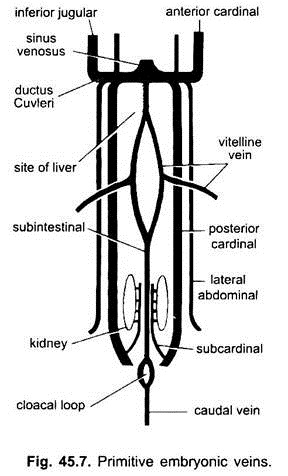
(iii) Se forman venas cardinales pareadas anteriores y posteriores, que transportan la sangre de la cabeza y las partes posteriores del cuerpo, respectivamente. Las venas cardinales anterior y posterior de cada lado se unen para formar un conducto Cuvieri o vena cardinal común que pasa hacia adentro a través del tabique transversal para ingresar al seno venoso.
En peces y salamandras (urodeles), una vena yugular inferior proviene del lado ventral de la cabeza para unirse a la vena cardinal común. No tiene homólogo en otros vertebrados. En un amniótico, un par de venas abdominales laterales o ventrales provienen de la pared corporal para ingresar a las venas cardinales comunes.
Piscis:
La vena cardinal común (conducto de Cuvier) entra en el seno venoso desde cada lado y está formada por la fusión de cardenales anteriores y posteriores. La sangre de la cabeza es recogida por los cardenales anteriores, y la sangre de los riñones y las gónadas es recogida por los cardenales posteriores.
También entran en los conductos de Cuvier las venas abdominales laterales pareadas que reciben sangre de la pared corporal y los apéndices pareados. El sistema portal renal está formado por la vena caudal y dos venas portales renales situadas laterales a los riñones que se capilarizaron en los riñones.
El sistema portal hepático transporta sangre desde el estómago y el intestino y la devuelve al hígado, donde, después de pasar por una serie de sinusoides, ingresa al seno venoso por medio de venas hapáticas pareadas.
En los teleosts, las venas abdominales laterales carecen de modo que la sangre de las subclavias, drenando los apéndices pectorales, entra directamente en el seno venoso, y la sangre de las venas ilíacas, drenando los apéndices pélvicos, pasa a los postcardinales.
En dipnoans, una sola vena abdominal ventral está presente, presumiblemente derivada de una fusión de abdominales laterales. Esta vena recibe sangre de los ilacs a través de venas pélvicas emparejadas y entra en el conducto derecho de Cuvier. Además, desde el sistema postcardinal derecho, una nueva vena, el postclaval hace su aparición que es de gran importancia en vertebrados superiores. Está conectado con la vena caudal y pasa hacia adelante a través del hígado hasta el seno.
En los embriones de los amniotas el lateral venas abdominales son conocidos como umbilical o alantoideo venas porque drenan el alantoides. Las venas vitelinas y umbilicales se pierden al nacer, solo persisten sus restos.
Modificaciones de venas:
Como en el caso de las arterias, las venas de diferentes vertebrados están dispuestas en el mismo plan fundamental. Cualquier variación mostrada por ellos sigue una secuencia en la serie de vertebrados. En su desarrollo, el sistema venoso de los vertebrados superiores pasa por las etapas observadas en embriones de formas inferiores.
1. Cuando se forma el hígado, la parte proximal de las venas vitelinas o venas subintestinales forma las venas hepáticas entre el hígado y el corazón, la parte distal de la vena vitelina izquierda o vena subintestinal se convierte en la vena porta hepática que forma sinusoides en el hígado para dar lugar a un sistema portal hepático presente en la lla.
2. Las venas cardinales anteriores persisten como venas yugulares internas.
3. Excepto en los peces, las venas cardinales comunes se convierten en venas precavales que entran en el seno venoso (anfibios, reptiles) o en la aurícula derecha (aves, mamíferos). En cada extremidad anterior se forma una vena subclavia que se une a la vena precaval.
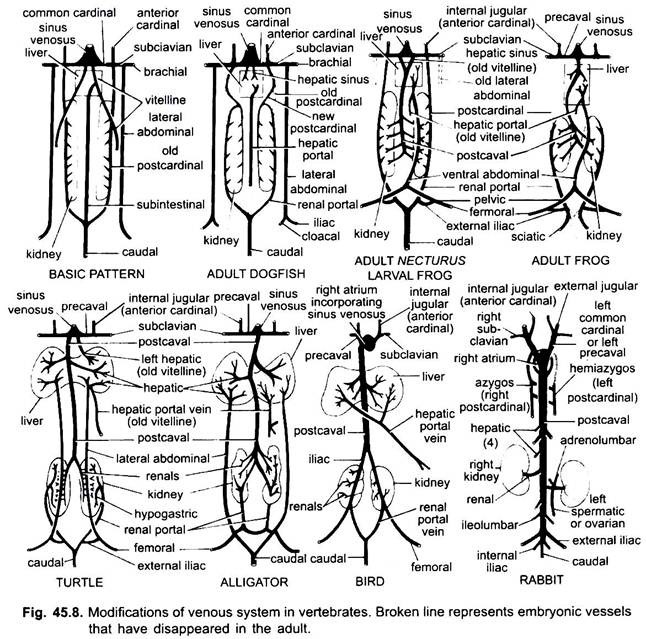
4. La vena caudal pierde sus conexiones con las venas subintestinales y subcardinales, su parte anterior se divide en dos ramas que se unen a las venas cardinales posteriores en todos, excepto en los mamíferos.
5. Las venas cardinales posteriores persisten como tales en peces, pero en otros, cada una se divide en dos porciones, una porción anterior que desaparece en anfibios, reptiles y aves, pero en mamíferos, la porción anterior derecha forma una vena ácigos, la izquierda puede formar una vena hemiácigos que pierde su conexión con la vena precaval y se une a la vena ácigos por una anastomosis transversal; las porciones posteriores de las venas cardinales posteriores que se han unido a la vena caudal, se convierten en las venas portales renales.
La vena porta renal de cada lado forma capilares en el riñón para formar un sistema portal renal que es completo en peces, anfibios y reptiles, pero se reduce mucho en las aves, y está ausente en los mamíferos porque las porciones posteriores de las venas cardinales posteriores desaparecen en los mamíferos.
6. Las venas portales renales de los peces recogen sangre solo de la cola, pero en anfibios, reptiles y aves también usurpan las venas de las patas.
7. En algunos peces y en todas las tetrápodos, las venas vitelinas se unen a la vena subcardinal para formar una vena postcaval. En anfibios y reptiles, la vena postcaval se extiende hasta los extremos posteriores de los riñones, en aves se une a las venas portales renales, reduciendo así el sistema portal renal, en mamíferos, la vena postcaval se une a las venas de las patas y la cola, de modo que el sistema portal renal se elimina por completo.
8. Las dos venas abdominales laterales persisten como tales en los peces, pero de Dipnoi hacia arriba se fusionan para formar una vena abdominal anterior que se une a la vena porta hepática cerca del hígado, la vena abdominal anterior, por lo tanto, conecta los sistemas portal renal y portal hepático. Se forma una vena ilíaca en cada extremidad posterior que se une a la vena abdominal anterior. La vena abdominal anterior comienza a perder importancia en reptiles, en aves se modifica para formar las venas epigástricas y coccigeo-mesentéricas, en mamíferos desaparece, excepto en Taquigloso.
9. En las formas de respiración aérea, las venas pulmonares salen de la aurícula izquierda y entran en los pulmones.
10. Las venas umbilicales (alantoicas) del embrión desaparecen cuando los pulmones se vuelven funcionales como órganos respiratorios.
Tetrapoda:
El sistema venoso de los anfibios se asemeja mucho al de los peces pulmonados, excepto que la vena abdominal ingresa al sistema portal hepático en lugar del seno venoso. Los cardenales anteriores persisten como venas yugulares internas en todos los tetrápodos adultos. Las venas yugulares inferiores están ausentes. Los cardenales comunes se convierten en la vena cava anterior o precavales, que se unen al seno venoso en anfibios y reptiles.
En aves y mamíferos, los precavales entran directamente en la aurícula derecha del corazón, porque el seno venoso está ausente. En hombre, gato, etc., el precaval izquierdo se pierde, por lo que la sangre del lado izquierdo entra en el precaval derecho a través de una rama braquiocefálica. En las salamandras sin pulmón, las venas pulmonares están ausentes ya que el tabique interauricular está incompleto.
Las venas post cardinales se unen a la vena caudal posteriormente y a las precavales anteriormente para formar los cardenales comunes en la rana larvaria y el Necturo.
En los reptiles, hay un mayor desarrollo de venas pulmonares y venas postcavas y una reducción en la importancia del sistema portal renal que lleva sangre a los riñones desde la parte posterior del cuerpo.
En las aves, hay dos venas precavales funcionales formadas por la unión de la yugular y la subclavia a cada lado, y un poste caval completo. Recibe sangre de las extremidades a través de los portales renales, que pasan a través de los riñones, pero no se rompen en capilares y no son comparables a los portales renales de los vertebrados inferiores.
En los mamíferos, puede haber una o dos venas precavales. Existe una única vena postcaval, cuyo desarrollo embriológico es muy complicado. Todas las venas cavales entran directamente en la aurícula derecha, ya que el seno venoso se absorbe en la pared de esta cámara del corazón en la vida embrionaria. No existe un sistema portal renal, aunque el sistema portal hepático se parece mucho al de otros vertebrados.