WERBUNG:
In diesem Artikel werden wir über das Verdauungssystem von Wirbeltieren mit Hilfe geeigneter Diagramme diskutieren.
Embryonaler Verdauungstrakt:
Archenteron:
Das embryonale Archenteron wird zur Auskleidung des adulten Verdauungstrakts und aller seiner Derivate. Das splanchnische Mesoderm fügt Bindegewebsschichten und glatte Muskeln um das Archenteron hinzu. Die ektodermale Invagination des Kopfes bildet das Stomodäum, das in die Mundhöhle führt, und eine ähnliche mittelventrale ektodermale Invagination bildet das Proctodaeum, das in den Hinterdarm führt.
ANZEIGEN:
Das Stomodäum wird die erwachsene Mundhöhle und führt zu Zahnschmelz, epitheliale Abdeckung der Zunge, Drüsen, z.B. Schleim, Gift und Speichel, etc., und Rathkes Beutel der Hypophyse anterior. Das Proctodaeum bildet entweder einen kleinen terminalen Teil der Kloake bei unteren Wirbeltieren und Rektum bei Säugetieren.
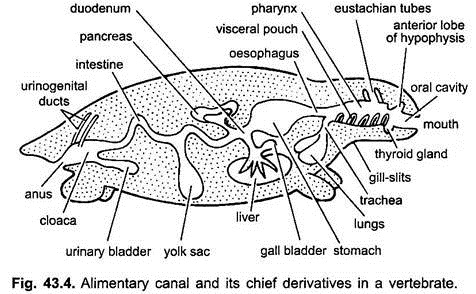
Der Verdauungskanal bei Embryonen vom Magen bis zur Kloake ist durch eine doppelte Peritoneumfalte, das sogenannte dorsale Mesenterium, an der dorsalen Körperwand und durch ein ventrales Mesenterium an der ventralen Körperwand befestigt. Bei Erwachsenen bleibt das dorsale Mesenterium bestehen, aber das ventrale Mesenterium verschwindet nur im Bereich der Leber und der Harnblase.
Verdauungstrakt des Erwachsenen:
Der Verdauungstrakt unterscheidet für verschiedene Funktionen in folgende Regionen – Mund, Mundhöhle, Rachen, Speiseröhre, Magen, Dünndarm, Dickdarm und Kloake. Folgende Auswüchse entstehen aus dem Verdauungstrakt- Munddrüsen, Rathkes Beutel, Schilddrüse, Kiemenspalten, Trommelfellhöhle, Thymus und andere Drüsen von Kiemenspalten, Luftröhre, Lunge, Schwimmblase, Leber, Bauchspeicheldrüse, Dottersack und Harnblase.
ANZEIGEN:
Histologie:
Die Wand des Verdauungskanals besteht aus vier konzentrischen Schichten.
Sie sind:
(i) Ein äußerstes viszerales Peritoneum oder seröser Mantel besteht aus Mesothelzellen und einer dünnen Bindegewebsschicht. Es fehlt in der Speiseröhre,
WERBUNG:
( ii) Darunter befindet sich eine Muskelschicht aus glatten Muskelfasern, die in äußeren longitudinalen und inneren kreisförmigen Muskelfasern angeordnet sind. Zwischen den beiden Muskelschichten befindet sich ein Netzwerk von Nervenzellen und Nervenfasern des autonomen Nervensystems, bekannt als Myenteric Plexus oder Plexus von Auerbach.
(iii) Unterhalb der Muskelschicht befindet sich eine Submukosa aus Bindegewebe mit elastischen Fasern, Fett-, Blut- und Lymphgefäßen, Nervenzellen und Faserndrüsen,
(iv) Die innerste Schicht ist eine Schleimhaut, die sich aus drei Regionen zusammensetzt:
(a) Äußerste-schmalste Muscularis-Schleimhaut aus äußeren longitudinalen und inneren kreisförmigen glatten Muskelfasern.
(b) Eine dünne Schicht Lamina propria aus Bindegewebe, Blutgefäßen, Nerven und Knötchen des lymphatischen Gewebes und
(c) Eine Basalmembran, die eine Schicht säulenförmiger Epithelzellen trägt, die häufig drüsig und bewimpert sind.
Mund:
Mund ist die Öffnung, die in die Mundhöhle führt. Bei Neunaugen (Cyclostomen) ist es eine kreisförmige Öffnung an der Basis des bukkalen Trichters und bleibt aufgrund fehlender Kiefer usw. dauerhaft offen. In Gnathostomen ist es terminal. Der Mund wird von Lippen begrenzt, die unbeweglich sind und bei Fischen, Amphibien und Reptilien aus verhornter Haut bestehen. Bei Säugetieren sind diese fleischig und muskulös.
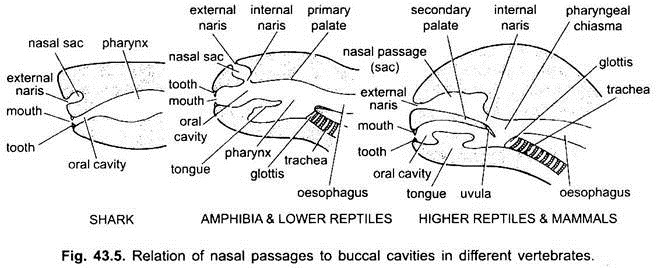
Mundhöhle:
WERBUNG:
Der Raum zwischen den Lippen und den Kiefern ist ein Vestibül. Es kann außen von Wangen und innen vom Zahnfleisch begrenzt sein. Schleimdrüsen der Wangen öffnen sich in den Vorraum. Der Mund öffnet sich in eine bukkale Höhle, die ein Raum zwischen dem Mund und dem Pharynx ist. Der genaue Punkt, an dem das stomodaeale Ektoderm und das pharyngeale Endoderm verschmelzen, ist variabel und nicht leicht zu erkennen.
Bei Elasmobranchen und den meisten Knochenfischen münden die Nasenhöhlen nicht in die Mundhöhle. Bei Chondrichthyes und Tetrapoden (Amphibien und die meisten Reptilien) öffnen sich die Nasenhöhlen durch Choanen oder innere Nasen, die primitiv anterior angeordnet sind, in die Mundhöhle, aber bei Krokodilen, Vögeln und Säugetieren werden sie aufgrund der Bildung eines sekundären Gaumens posterior im Pharynx, der den Nasengang der Atemwege effektiv von der Mundhöhle oder dem Nahrungsdurchgang trennt.
Bei Vögeln ist dieser Gaumen gespalten, wodurch Nasen- und Mundhöhlen miteinander kommunizieren. Bei Säugetieren wird der sekundäre Gaumen posterior als membranöser weicher Gaumen fortgesetzt. Beim Menschen weichen Gaumen hängt in den Kehlkopf Rachen in Form von fleischigen Prozess, genannt Uvula.
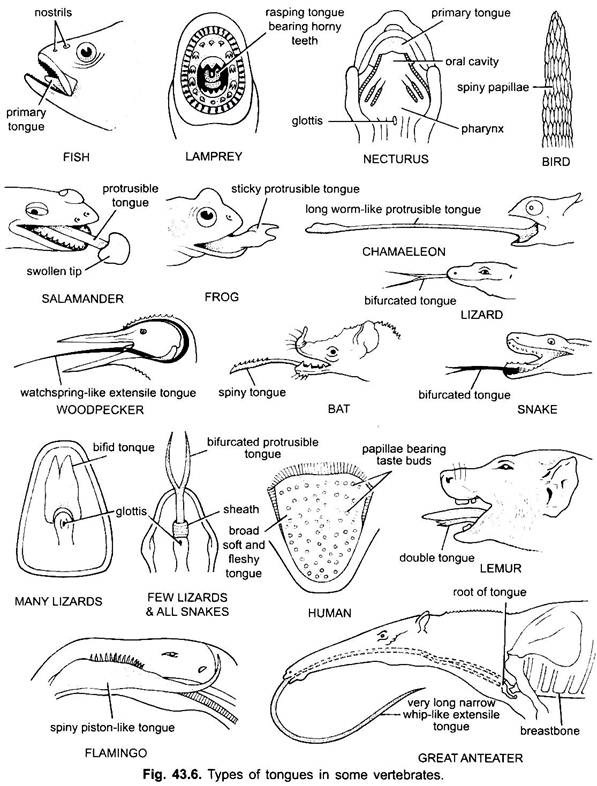
Derivate der Mundhöhle:
1. Munddrüsen:
Es gibt zwei Arten von integumetären mehrzelligen Drüsen, die sich in die Mundhöhle öffnen. Sie sind Schleimdrüsen und enzymatische Drüsen. Fische und Wasseramphibien haben nur Schleimdrüsen. Reptilien haben Drüsen in Gruppen, wie Gaumen-, Lingual-, Sublingual- und Labialdrüsen, die je nach Standort benannt sind.
Bei Giftschlangen sind die oberen Labialdrüsen so modifiziert, dass sie Gift absondern, während bei den Gilamonstern die sublingualen Drüsen Gift produzieren. Vögel haben sublinguale Drüsen und eine Drüse im Mundwinkel. Säugetiere haben viele kleine Schleimdrüsen neben echten und vergrößerten Speicheldrüsen, die enzymatisch sind. Sie sind parotide, sublinguale, submaxilläre und infraorbitale Speicheldrüsen, die Mucin und Ptyalin absondern.
2. Zunge:
Die Zunge kommt vor allem bei allen Wirbeltieren vor. Zunge in Wirbeltieren zeigen viel Vielfalt und sind nicht homolog. In Cyclostomen gibt es eine muskulöse, fleischige, raspelnde Zunge mit geilen Zähnen zum Raspeln der Haut und Muskeln ihrer Beute.
Fische haben eine primäre Zunge, die aus einer fleischigen Falte des bukkalen Bodens besteht. Es hat keine Muskeln, aber Rezeptoren und Zähne sind bei einigen Knochenfischen auf der Zunge vorhanden. Die Zunge ist mit Schleimhaut bedeckt. Bei einigen Amphibien ist die Zunge entweder fehlt oder unbeweglich. Die meisten Amphibien haben jedoch eine hervorstehende Zunge, und bei einigen Fröschen und Kröten kann sie bei Nichtgebrauch auf sich selbst zurückgefaltet werden.
Sie kann durch schnellen Lymphfluss aus dem Mund geworfen werden, um die Insekten einzufangen. Die Zunge bei Eidechsen und Schlangen ist oft hoch entwickelt. In Chamäleons ist es sehr erweiterbar, um Insekten zu fangen. Die Spitze ist verdickt und klebrig. Die gegabelte Zungenspitze in Schlangen dient dazu, chemische Reize von der äußeren Umgebung auf die gepaarten vomero-nasalen Organe auf dem Dach des Mundes zu übertragen.
Bei Schildkröten und Krokodilen kann die Zunge nicht verlängert werden. Die Amniotenzunge hat freiwillige Muskeln, sie empfängt den Hypoglossusnerv und hat Drüsen und Geschmacksknospen. Es entwickelt auch intrinsische Muskeln, die die Zunge bewegen. Bei Vögeln ist die Zunge schlank und hat eine geile Hülle. Bei einigen Vögeln ist die Zunge unbeweglich, während sie bei einigen Vögeln lang, langwierig und oft zum Einfangen des Futters verwendet wird.
Bei den meisten Säugetieren, mit Ausnahme von Walen, ist die Zunge hoch entwickelt und aufgrund der Anwesenheit einer Reihe von intrinsischen Muskeln in der Lage, sich zusätzlich zur Extension und Retraktion beträchtlich zu bewegen.
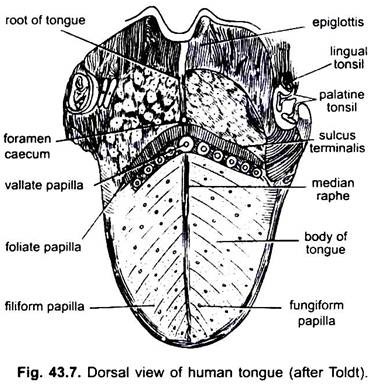
Bei Säugetieren bildet die Schleimhaut unterhalb der Zunge eine mediane Falte, Frenulum genannt, die die Zunge mit dem Mundboden verbindet. Bei Säugetieren trägt die obere Oberfläche der Zunge vier Arten von Papillen (filiform, fungiform, foliate und circumvallate), die Geschmacksknospen außer fadenförmigen Papillen tragen.
3. Zähne:
Wirbeltiere haben zwei Arten von Zähnen, die an Kieferknochen befestigt sind – epidermale Zähne und echte Zähne. Epidermale Zähne werden am besten in Cyclostomen entwickelt. Sie sind harte, konische, hornartige Strukturen, die aus dem Stratum corneum stammen. Bei Neunaugen finden sie sich an den Wänden des bukkalen Trichters und auf der Zunge. Kaulquappenlarven von Fröschen und Kröten haben gezahnte Epidermiszähne in Reihen auf den Lippen. Bei Säugetieren hat das erwachsene Entenschnabeltier epidermale Zähne.
Wahre Zähne:
Zähne kommen bei Bartenwalen und Ameisenbären bei Säugetieren sowie bei Agnathanen, Stören, einigen Kröten, Sireniern, Schildkröten und modernen Vögeln usw. nicht vor. Bei niederen Wirbeltieren (wie Fischen, Amphibien und den meisten Reptilien) können Zähne auf unbestimmte Zeit ersetzt werden, solche Zähne werden Polyphyodont genannt. Diese Zähne sind homodont (ähnlicher Typ) und Acrodont. (mit den Kieferknochen).
Bei den meisten Säugetieren sind die Zähne Diphyodont, Thecodont und Heterodont. Bei einigen Säugetieren sind dies Monophyodont mit nur einem Satz Zähne, z. B. Maulwürfe, indisches Eichhörnchen. Die Zähne ähneln in ihrer Struktur den placoiden Schuppen von Haien, die aus einer zentralen Pulpahöhle bestehen, um die herum eine dicke, aber weiche Schicht vorhanden ist, das Dentin, das äußerlich von einem dünnen, extrem harten Zahnschmelz bedeckt ist. Diese sollen von knöchernen Schuppen von Ostracoderms und Placoderms abgeleitet haben. Für Details können Leser das Gebiss bei Säugetieren sehen.
4. Adenohypophyse:
Der Hypophysenvorderlappen entwickelt sich als dorsale Evagination des Stomodäums, Rathkes Beutel genannt, der sich zu den Hypophysenvorder- und Mittellappen verengt (Adenohypophyse). Der hintere Lappen der Hypophyse oder Neurohypophyse ist die ventrale Evagination des Diencephalons, genannt Infundibulum. Somit ist es nervöser Teil.
Pharynx:
Der Teil des Verdauungskanals unmittelbar hinter der Mundhöhle ist ein mit Endoderm ausgekleideter Pharynx. Es ist eine gemeinsame Passage, die sowohl der Verdauung als auch der Atmung dient. Als Teil des Verdauungssystems wird es als Durchgang für Nahrung von der Mundhöhle zur Speiseröhre verwendet, seine Muskeln initiieren das Schlucken.
Bei Fischen ist der Pharynx groß und seitlich für Kiemenschlitze perforiert, während er bei Tetrapoden kurz ist und Öffnungen von Nasenlöchern trägt. Bei Embryonen gibt die Wand des Pharynx eine Reihe von Evaginationen ab, die sich zu Spirakeln, Kiemenspalten, Luftblasen, Lungen, Mandeln und einigen endokrinen Drüsen (z. B. Thymus, Schilddrüse und Nebenschilddrüse) entwickeln.
Speiseröhre:
Die Speiseröhre ist bei den meisten Fischen und Amphibien kurz, weil ihnen der Hals fehlt, aber bei Amnioten ist sie aufgrund des Vorhandenseins des Halses länger. Die Speiseröhre von Reptilien ist länger als die von Fischen und Amphibien. Bei körner- und fleischfressenden Vögeln wird ein Teil der Speiseröhre zu einem sackartigen Beutel namens Crop vergrößert, der zur Aufbewahrung von schnell gefressenen Nahrungsmitteln dient.
Der Ernte fehlen im Wesentlichen Verdauungsdrüsen obwohl die Ernte bei Tauben bei beiden Geschlechtern 2 Fruchtdrüsen aufweist, handelt es sich in Wirklichkeit nicht um Drüsen, sondern um zellbildende Strukturen, die Zellen bilden ‚Taubenmilch‘, die an die Jungen verfüttert wird. Bei Säugetieren ist die Speiseröhre lang, fehlt Drüsen und variiert in Bezug auf die Länge des Halses.
Es geht durch das Zwerchfell, der Teil unterhalb des Zwerchfells ist mit viszeralem Peritoneum bedeckt, das vom oberen Teil fehlt. Die Speiseröhre hat Schleimdrüsen. Seine Auskleidung bildet Längsfalten oder fingerartige fleischige Papillen (Elasmobranchen) oder Hornpapillen bei Meeresschildkröten.
Histologisch unterscheidet sich die Speiseröhre vom Rest des Verdauungskanals in drei Punkten:
(i) Sie hat kein viszerales Peritoneum, da sie außerhalb des Coeloms liegt, ihre äußerste Deckschicht ist eine dünne Tunica adventitia.
(ii) Die Muskelfasern im vorderen Teil sind gestreift, der mittlere Teil hat sowohl gestreifte als auch ungestreifte Muskeln und der hintere Teil hat nur ungestreifte Muskeln. Aber es gibt Ausnahmen bei wiederkäuenden Säugetieren, alle Muskeln sind gestreift oder freiwillig.
(iii) Die Schleimhaut besteht aus geschichteten Plattenepithelzellen und nicht aus Säulenzellen.
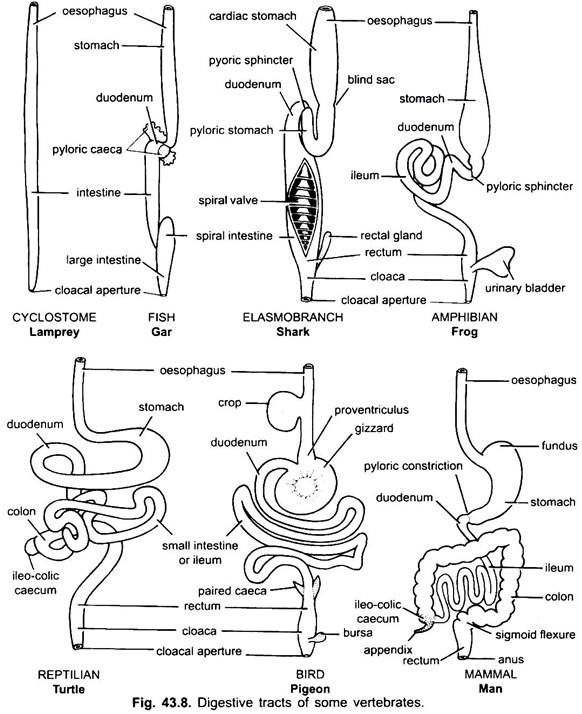
Magen:
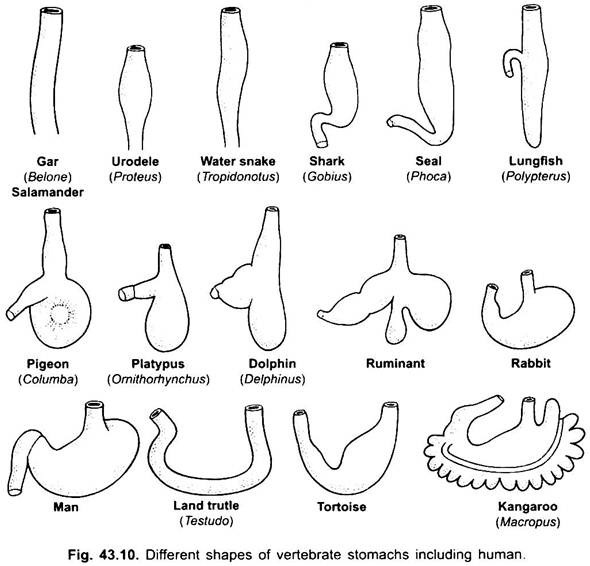
Bei Zyklostomen, Chimären, Lungenfischen und einigen primitiven Teleostfischen gibt es praktisch keinen Magen, da er keine Magendrüsen hat, aber bei den meisten Fischen und Tetrapoden ist er zur Lagerung und Mazeration fester Nahrung und zur Verdauung von Nahrung erweitert, da er Magendrüsen enthält.
Der erste Teil des Magens neben der Speiseröhre ist die Herzregion und das untere Ende in der Nähe des Darms ist die Pylorusregion, die eine Pylorus- oder Pylorusklappe aufweist, in der die Schleimhaut von einem dicken Schließmuskel umgeben ist, der das Öffnen und Schließen des Pylorusmagens in den Darm reguliert.
Magen ist gerade in Cyclostomen, GAR, Belone, etc., und spindelförmig in Proteus, Necturus, einigen Eidechsen und Schlangen. Bei Schildkröten und Schildkröten ist es eine breite gebogene Röhre, und bei Elasmobranchen ist der Magen J-förmig. Bei Krokodilen und Vögeln besteht der Magen aus zwei Teilen, einem Proventriculus mit Magendrüsen und einem stark muskulösen Muskelmagen, der die Pylorusregion darstellt und eine harte, verhornte Auskleidung zum Mahlen von Nahrungsmitteln aufweist.
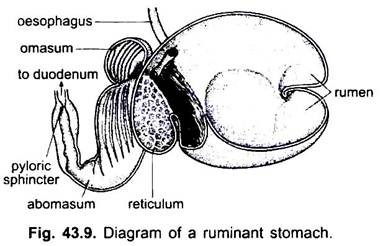
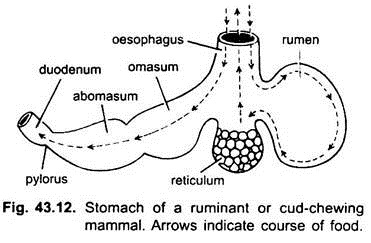
Bei Säugetieren liegt der Magen quer und kann ein einfacher Sack sein oder in 3 Regionen unterteilt sein, nämlich Herz, Fundus und Pylorus, und jede Region hat ihre Magendrüsen. Bei vielen Wiederkäuern hat der Magen vier Kammern – einen Pansen, ein Retikulum, ein Omasum und ein Abomasum. Es wird behauptet, dass die ersten drei Kammern Modifikationen der Speiseröhre sind, und Abomasum ist der wahre Magen, der die Herz-, Fundus- und Pylorus-Teile des Magens darstellt.
Es wurde embryologisch gezeigt, dass alle vier Kammern modifizierte Regionen des Magens sind. In Kamelen gibt es kein Omasum, der Pansen und das Retikulum haben beutelartige Wasserzellen, von denen einst angenommen wurde, dass sie Wasser speichern, aber sie sind wahrscheinlich verdauungsfördernd.
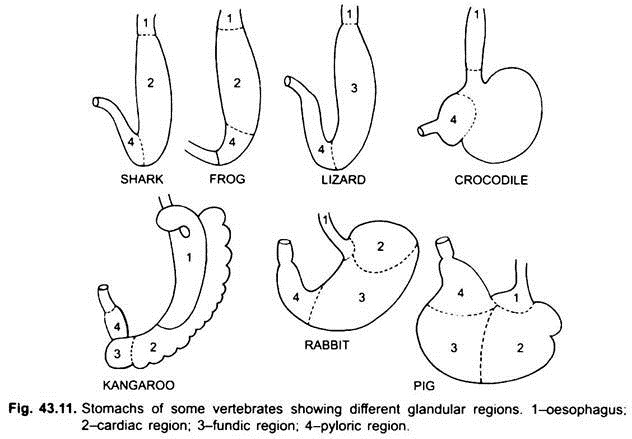
Histologisch weist der Magen die typischen Teile des Verdauungskanals auf, weist jedoch zwei Besonderheiten auf: Die Muscularis mucosa besteht aus einer äußeren Längsschicht und einer inneren kreisförmigen Muskelschicht. Die Epithelauskleidung ist dick mit verschiedenen Arten von Drüsenzellen, die Magendrüsen von drei Arten bilden, die als Herz-, Fundus- und Pylorus-Magendrüsen bezeichnet werden.
Die Herz- und Pylorusdrüsen scheiden nur Schleim aus ihren Oberflächenzellen aus. Fundische Drüsen (oder Herzdrüsen in einigen) haben drei Arten von Zellen, Schleimhalszellen produzieren Schleim, oxyntische Zellen produzieren Salzsäure, sie können auch in der Herzregion vorhanden sein, Zymogenzellen oder peptische Zellen sezernieren Pepsin.
Bei den meisten Tieren sezernieren die Zymogenzellen auch zwei Proenzyme, Propepsin und Prorennin, die durch Salzsäure in Pepsin bzw. Die Sekrete aller Magenzellen bilden eine Mischung namens Magensaft.
Dünndarm:
Der Dünndarm ist ein langer, schmaler und gewundener Schlauch nach dem Pylorus. Es ist der wichtigste Teil des Verdauungstraktes, da die Verdauung und Aufnahme von Nahrung darin stattfindet. In Cyclostomen ist der Darm eine kurze gerade Röhre mit einem spiralförmig angeordneten Längslappen, der sich hinein erstreckt.
In Elasmobranchen ist es in kleine und große Portionen unterteilt, und der kleine Teil hat ein Spiralventil, das die Absorptionsfläche stark vergrößert. Eine Spiralklappe ist auch im Dünndarm einiger primitiverer Knochenfische vorhanden, fehlt jedoch in höheren Formen, in denen der Darm lang und gewunden ist.
Bei Caecilianern ist es wenig gewunden und nicht in einen kleinen und großen Trakt differenziert. Bei Fröschen und Kröten ist es relativ lang und gewunden. Bei Reptilien ist es stärker gewickelt als bei Amphibien. Zum ersten Mal bei Wirbeltieren entsteht an der Kreuzung von Dünn- und Dickdarm ein blindes Divertikel oder blindes Divertikel.
Dies ist jedoch nicht bei allen Reptilien dauerhaft. Bei Vögeln ist der Dünndarm gewickelt oder geschlungen und ein oder zwei Koliken caeca sind auch an der Kreuzung von Dünn- und Dickdarm vorhanden. Bei den meisten Säugetieren ist auch der Dünndarm proportional lang und gewunden. Seine Länge korreliert jedoch mit den Ernährungsgewohnheiten. Bei Pflanzenfressern ist es im Vergleich zu Insektenfressern und Fleischfressern relativ länger.
An der Kreuzung von Dickdarm und Dünndarm befindet sich eine blinde Tasche oder ein blinder Blinddarm, der bei fleischfressenden Arten im Allgemeinen klein und bei vielen Pflanzenfressern ziemlich lang ist. Der erste Teil des Dünndarms ist der Zwölffingerdarm, der vom Pylorus aus kurz ist und über den Eingang der Pankreas- und Lebergänge hinaus endet.
Es hat viele gefaltete Zotten und enthält verzweigte Brunner-Drüsen in der Submukosa, die Schleim, etwas alkalische wässrige Flüssigkeit und ein wenig Enzym absondern. Der Zwölffingerdarm produziert auch zwei Hormone namens Sekretin und Cholecystokinin, die die Bauchspeicheldrüse und die Gallenblase stimulieren, um ihre Säfte freizusetzen. Kanäle aus der Gallenblase und der Bauchspeicheldrüse münden in den Zwölffingerdarm.
Hinter dem Zwölffingerdarm befindet sich ein Ileum, das nur bei Säugetieren in ein vorderes kleineres Jejunum und ein hinteres längeres Ileum unterschieden wird. Eine große Anzahl kleiner Verdauungsdrüsen ist im Dünndarm vorhanden. Sie sind röhrenförmige Drüsen oder Krypten von Lieberkuhn, die über die gesamte Länge gefunden werden, sie scheiden Schleim und einen Succus entericus aus, der mehrere Enzyme enthält.
Die Auskleidung des Dünndarms wird zu kleinen Zotten gefaltet, die die Oberfläche für die Sekretion und Absorption vergrößern. Die Zotten sind dicht von winzigen fingerartigen Vorsprüngen bedeckt, die als Mikrovilli bezeichnet werden und die Absorption in die Zotten unterstützen. Bei Säugetieren befinden sich Knötchen von lymphatischem Gewebe, die Peyer-Flecken genannt werden, auf dem Ileum.
Dickdarm:
Der Dickdarm hat einen größeren Durchmesser als der Dünndarm. Bei Fischen, Amphibien, Reptilien und Vögeln ist es im Allgemeinen kurz, bei Säugetieren jedoch lang. In niedrigeren Formen bildet der Dickdarm ein Rektum, aber in Tetrapoden hat er einen Dickdarm und ein terminales Rektum. Bei den meisten Fischen und Amphibien führt der terminale Teil des Mastdarms in eine Kloake, die vom Proctodaeum gebildet wird.
Das Rektum, die Ausscheidungsgänge und die Genitalgänge münden in die Kloake und öffnen sich durch eine Kloakenöffnung nach außen. Aber bei vielen Knochenfischen und allen Säugetieren (außer Monotremen) haben das Rektum und die Urinogenitalgänge separate Öffnungen nach außen; die Öffnung des ersteren ist ein Anus.
Das Rektum von Säugetieren ist nicht homolog zum Rektum von Wirbeltieren, da es bei Säugetieren durch Partitionierung embryonaler Kloaken gewonnen wird. Bei den meisten Wirbeltierembryonen gibt es einen postanalen Darm als Verlängerung des Darms in den Schwanz, der jedoch später verschwindet.
Bei Elasmobranchen trägt der Dickdarm ein Paar Rektaldrüsen, die Schleim und Natriumchlorid absondern. Bei Amnioten befindet sich an der Verbindung von Dünn- und Dickdarm eine Ileokolikklappe, die bei Fischen fehlt. Es verhindert, dass Bakterien aus dem Dickdarm in das Ileum gelangen.
Bei Amnioten entsteht aus dieser Kreuzung ein ileokolischer Blinddarm, der bei Vögeln zwei ist. Es enthält Zellulose verdauende Bakterien. Es ist sehr lang in pflanzenfressenden Säugetieren (Kaninchen, Pferd, Kuh, etc.). Bei Primaten ist der Blinddarm klein und hat einen verkümmerten Wurmfortsatz.
Verdauungsdrüsen:
1. Leber:
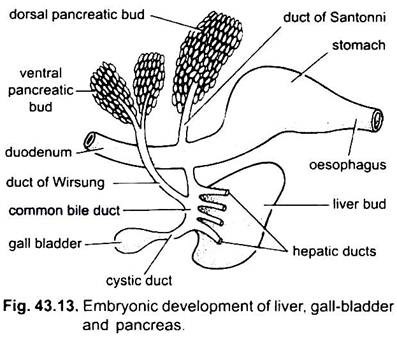
Die Leber entsteht als einfacher oder doppelter Auswuchs aus der Bauchwand des embryonalen Archenterons. Dieses Auswuchs bildet ein hohles Leberdivertikel, das sich bald in einen vorderen Teil differenziert, der sich zur Leber und ihren Gallengängen vermehrt, und einen hinteren Teil, aus dem die Gallenblase und der Zystengang entstehen. Die Gallengänge verbinden sich zu einem Lebergang, der sich mit dem Ductus cysticus zu einem gemeinsamen Gallengang oder Ductus choledochus verbindet. Die Region des Archenterons, aus der die Leber entsteht, wird zum Zwölffingerdarm.
Die Leber ist die größte gelappte Drüse im Körper, die durch eine doppelte Schicht Peritoneum vom Querseptum oder seinem Vertreter suspendiert ist.
Eine Gallenblase dient zur Speicherung der von den Leberzellen abgesonderten Galle, liegt in der Leber und fließt durch den gemeinsamen Gallengang, der durch die Vereinigung von Ductus cysticus und Ductus hepaticus gebildet wird, in den Zwölffingerdarm. Eine Gallenblase ist nicht unverzichtbar und fehlt vielen Vögeln und Säugetieren.
Eine Leber ist in allen Wirbeltieren vorhanden. In Cyclostomen ist es klein, einlappig (Neunaugen) und zweilappig in Hagfischen. Es ist zweilappig in Elasmobranchen, zwei oder drei lappig in Knochenfischen, Amphibien, Reptilien und Vögeln und viele lappig in Säugetieren. Leber ist lang, schmal und zylindrisch in Fischen, Urodelen und Schlangen.
Es ist kurz, breit und abgeflacht bei Vögeln und Säugetieren. Eine Gallenblase und ein Gallengang sind in Larvenzyklostomen vorhanden, aber beim Erwachsenen fehlen sie. Fische, Amphibien und Reptilien haben im Allgemeinen eine Gallenblase, die jedoch bei vielen Vögeln fehlt. Die meisten Säugetiere haben eine Gallenblase, aber es fehlt in Cetacea und Ungulata.
Die Leber scheidet eine wässrige, alkalische Galle aus, hat aber keine Enzyme. Es neutralisiert den Säuregehalt von Lebensmitteln, die in den Zwölffingerdarm gelangen. Es hilft bei der Verdauung von Fetten.
2. Pankreas:
Pankreas wird aus dem Endoderm des embryonalen Archenterons gebildet. Ein einzelnes dorsales Divertikel aus dem embryonalen Zwölffingerdarm und ein oder zwei ventrale Auswüchse aus der Leber bilden Pankreasdivertikel. Die proximalen Teile der Divertikel bilden Pankreasgänge, aber diese Kanäle unterliegen einer Reduktion oder Fusion, so dass nur ein oder zwei Pankreasgänge im Erwachsenen verbleiben, sie öffnen sich entweder getrennt oder nach Vereinigung mit dem gemeinsamen Gallengang in den Zwölffingerdarm.
Die distalen Teile der Divertikel werden knospend, um die Hauptmasse der Pankreaszellen zu bilden, denen mesodermale Derivate zugesetzt werden. Somit wird eine einzelne Drüse hergestellt, die mehrere Lappen aufweist, die entweder eine diffuse oder eine kompakte Bauchspeicheldrüse bilden.
Die Bauchspeicheldrüse ist sowohl eine exokrine als auch eine endokrine Drüse, die durch empfindliche Bindegewebsstränge miteinander verbunden ist. Der exokrine Teil sondert Verdauungsenzyme ab, die durch Pankreasgänge in den Zwölffingerdarm gegossen werden. Während der endokrine Teil Hormone wie Insulin und Glucagon absondert.
Eine Bauchspeicheldrüse ist in allen Wirbeltieren vorhanden. Bei Neunaugen, einigen Knochenfischen, Lungenfischen und unteren Tetrapoden ist es ein diffuses Organ, das in Leber, Mesenterien und Darmwand eingebettet ist. Hagfische haben eine kleine Bauchspeicheldrüse. Elasmobranchs haben eine gut definierte bilobed Bauchspeicheldrüse. In höheren Tetrapoden ist es im Allgemeinen eine kompakte Drüse. Ein oder zwei Pankreasgänge öffnen sich in den Zwölffingerdarm.