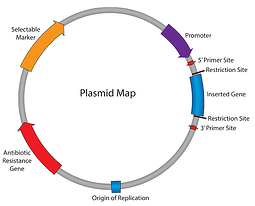
 Bisher haben wir uns in unserer Plasmidserie 101 durch die Plasmidkarte gearbeitet: Antibiotikaresistenz, Replikationsursprung und so weiter. Bis zu diesem Punkt können wir unser Plasmid replizieren und sicherstellen, dass die Zellen es beibehalten; Der nächste Schritt besteht darin, das Plasmid dazu zu bringen, unser interessierendes Gen zu exprimieren. Geben Sie den Promotor ein – das Element, das für die Initiierung der Transkription Ihres Inserts in RNA verantwortlich ist.
Bisher haben wir uns in unserer Plasmidserie 101 durch die Plasmidkarte gearbeitet: Antibiotikaresistenz, Replikationsursprung und so weiter. Bis zu diesem Punkt können wir unser Plasmid replizieren und sicherstellen, dass die Zellen es beibehalten; Der nächste Schritt besteht darin, das Plasmid dazu zu bringen, unser interessierendes Gen zu exprimieren. Geben Sie den Promotor ein – das Element, das für die Initiierung der Transkription Ihres Inserts in RNA verantwortlich ist.
In der Praxis beschreibt der Begriff „Promotor“ die Kombination von Promotor (RNA-Polymerase-Bindungsstelle) und Operatoren (Response-Elemente). Promotoren sind etwa 100 bis 1000 Basenpaare lang und befinden sich stromaufwärts ihrer Zielgene. Die Sequenz der Promotorregion steuert die Bindung der RNA-Polymerase und der Transkriptionsfaktoren, daher spielen Promotoren eine große Rolle bei der Bestimmung, wo und wann Ihr interessierendes Gen exprimiert wird.
Die RNA-Polymerase(n)
RNA wird aus DNA mittels einer RNA-Polymerase (RNAP) transkribiert. In Bakterien geschieht dies durch ein einzelnes Enzym; Eukaryoten haben jedoch mehrere Polymerasen, die jeweils für eine bestimmte Teilmenge von RNAs verantwortlich sind. Um diese Spezifität zu erlangen, kann der eukaryotische RNAP spezifische Promotorelemente erkennen und an diese binden. Dies bedeutet, dass der Promotor, der in Ihrem Plasmid-Backbone vorhanden ist, mit der Art der RNA kompatibel sein muss, die hergestellt werden muss: Wenn Sie mRNA (für die Genexpression) möchten, müssen Sie einen RNAP II-Promotor verwenden, während kleine RNAs (wie shRNA) von den RNAP III-Promotoren transkribiert werden. Dieser Beitrag enthält Promotoren für die allgemeine RNAP II- und RNAP III-Transkription; Die Verwendung viraler LTRs als RNAP II-Promotoren wird jedoch häufig in lentiviralen und retroviralen Konstrukten eingesetzt, und wir werden diese in einem zukünftigen Beitrag zu viralen Vektorteilen diskutieren.
Promotorspezifität
Neben der Auswahl eines Promotors basierend auf dem Typ des RNA-Transkripts müssen Sie auch sicherstellen, dass Ihr Plasmid einen Promotor hat, der für die Arbeit in Ihrem Wirtsorganismus geeignet ist. Da sich die Transkriptionsmaschinerie zwischen Zelltypen oder Organismen unterscheidet, müssen die Promotoren ähnlich variabel sein. Bakterielle Promotoren wirken nur in prokaryotischen Zellen und typischerweise nur in derselben oder eng verwandten Spezies, von der sie abgeleitet wurden. In ähnlicher Weise erfordern die verschiedenen eukaryotischen Zelltypen (Säugetiere, Hefen, Pflanzen usw.) einzigartige Promotoren und es gibt sehr wenig Crossover. Im Allgemeinen sind Promotoren in Bakterien weniger vielfältig und komplex und haben weniger Teile als solche in eukaryotischen Zellen. Einige Promotoren sind konstitutionell aktiv und ständig aktiv, während andere sorgfältiger kontrolliert werden. Regulierte Promotoren können nur in bestimmten Geweben oder zu bestimmten Zeiten in der Entwicklung wirken, oder es kann Möglichkeiten geben, sie nach Belieben mit einer Chemikalie, Wärme oder Licht ein- oder auszuschalten. In der Zelle werden Promotoren selbst durch noch andere regulatorische Faktoren gesteuert: Enhancer, Grenzelemente, Isolatoren und Schalldämpfer; Es kann jedoch zu einer gewissen „Undichtigkeit“ der Transkription kommen. Dies ist normalerweise kein großes Problem für Zellen, aber es kann Forschungsergebnisse verwirren oder sogar Ihre Zellen töten, wenn Ihr interessierendes Gen toxisch ist. Um dies zu bekämpfen, haben Wissenschaftler synthetische Promotoren entwickelt, die typischerweise eine Kombination anderer Promotorelemente enthalten und tendenziell strenger reguliert sind.
Gemeinsame Promotoren für Eukaryoten und Prokaryoten
Wir haben unten zwei Referenztabellen eingefügt, in denen einige der häufigsten Bakterien- und Säugetierpromotoren aufgeführt sind. Diese Listen sind keineswegs erschöpfend, sollten aber ein guter Anfang sein, wenn Sie versuchen, Ihren perfekten Promoter auszuwählen!
Eukaryotische Promotoren
| Promotor | Primär verwendet für | RNA-Transkript | Beschreibung | Expression | Zusätzliche Überlegungen |
| CMV | Allgemeine Expression | mRNA | Starker Säugetier-Expressionspromotor aus dem humanen Cytomegalovirus | Konstitutiv | Kann eine Enhancer-Region enthalten. Kann in einigen Zelltypen zum Schweigen gebracht werden. |
| EF1a | Allgemeine Expression | mRNA | Starke Säugetierexpression aus menschlichem Elongationsfaktor 1 alpha | Konstitutiv | Neigt unabhängig vom Zelltyp oder der Physiologie zu einer konsistenten Expression. |
| SV40 | Allgemeine Expression | mRNA | Säugetier-Expressionspromotor aus dem Simian vacuolating Virus 40 | Konstitutiv | Kann einen Enhancer enthalten. |
| PGK1 (Mensch oder Maus) | Allgemeine Expression | mRNA | Säugetierpromotor aus Phosphoglyceratkinase-Gen. | Konstitutiv | Weit verbreitete Expression, kann aber je nach Zelltyp variieren. Neigt dazu, der Promotor-Down-Regulation aufgrund von Methylierung oder Deacetylierung zu widerstehen. |
| Ubc | Allgemeine Expression | mRNA | Säugetierpromotor aus dem humanen Ubiquitin C-Gen | Konstitutiv | Wie der Name schon sagt, ist dieser Promotor ubiquitär. |
| menschliches Beta-Aktin | Allgemeine Expression | mRNA | Säugetier-Promotor aus dem Beta-Aktin-Gen | Konstitutiv | Ubiquitär. Hühnerversion wird häufig in Promotorhybriden verwendet. |
| CAG | Allgemeine Expression | mRNA | Starker hybrider Säugetierpromotor | Konstitutiv | Enthält CMV-Enhancer, Hühner-Beta-Aktin-Promotor und Kaninchen-Beta-Globin-Spleißakzeptor. |
| TRE | Allgemeine Expression | mRNA | Tetracyclin-Antwortelement-Promotor | Induzierbar mit Tetracylin oder seinen Derivaten. | enthält typischerweise einen minimalen Promotor mit geringer basaler Aktivität und mehrere Tetracyclin-Operatoren. Die Transkription kann ein- oder ausgeschaltet werden, je nachdem, welcher tet-Transaktivator verwendet wird. |
| UAS | Allgemeine Expression | mRNA | Drosophila-Promotor mit Gal4-Bindungsstellen | Spezifisch | Erfordert die Anwesenheit des Gal4-Gens, um den Promotor zu aktivieren. |
| Ac5 | Allgemeine Expression | mRNA | Starker Insektenpromotor aus dem Drosophila Actin 5c-Gen | Konstitutiv | Häufig verwendet in Expressionssystemen für Drosophila. |
| Polyedrin | Allgemeine Expression | mRNA | Starker Insektenpromotor aus Baculovirus | Konstitutiv | Häufig verwendet in Expressionssystemen für Insektenzellen. |
| CaMKIIa | Genexpression für die Optogenetik | mRNA | Ca2 + / Calmodulin-abhängiger Proteinkinase-II-Promotor | Spezifisch | Zur neuronalen / ZNS-Expression verwendet. Moduliert durch Calcium und Calmodulin. |
| GAL1, 10 | Allgemeine Expression | mRNA | Hefe benachbarte, divergent transkribierte Promotoren | Induzierbar mit Galactose; repressibel mit Glucose | Können unabhängig oder zusammen verwendet werden. Geregelt durch GAL4 und GAL 80. |
| TEF1 | Allgemeine Expression | mRNA | Hefe-Transkriptions-Elongationsfaktor-Promotor | Konstitutiv | Analog zum EF1A-Promotor von Säugetieren. |
| GDS | Allgemeine Expression | mRNA | Starker Hefe-Expressionspromotor aus Glyceraldehyd-3-Phosphage-Dehydrogenase | Konstitutiv | Sehr stark, auch TDH3 oder GAPDH genannt. |
| ADH1 | Allgemeine Expression | mRNA | Hefepromotor für Alkoholdehydrogenase I | Unterdrückt durch Ethanol | Version in voller Länge ist stark mit hoher Expression. Abgeschnittene Promotoren sind konstitutiv mit geringerer Expression. |
| CaMV35S | Allgemeine Expression | mRNA | Starker Pflanzenpromotor aus dem Blumenkohlmosaikvirus | Konstitutiv | Aktiv in dikotylen, weniger aktiv in monokotylen, mit einer gewissen Aktivität in tierischen Zellen. |
| Ubi | Allgemeine Expression | mRNA | Pflanzenpromotor aus Mais-Ubiquitin-Gen | Konstitutiv | Ergibt eine hohe Expression in Pflanzen. |
| H1 | kleine RNA-Expression | shRNA | Aus dem humanen Polymerase-III-RNA-Promotor | Konstitutiv | Kann eine etwas geringere Expression als U6 aufweisen. Kann eine bessere Expression in neuronalen Zellen haben. |
| U6 | kleine RNA-Expression | shRNA | Vom humanen U6 kleiner Kernpromotor | Konstitutiv | Murines U6 wird ebenfalls verwendet, kann jedoch weniger effizient sein. |
Prokaryotische Promotoren
| Promotor | Primär verwendet für | Beschreibung | Expression | Zusätzliche Überlegungen |
| T7 | in vitro Transkription/ allgemeine Expression | Promotor aus T7-Bakteriophagen | Konstitutiv, erfordert jedoch T7-RNA-Polymerase. | Bei Verwendung für die In-vitro-Transkription steuert der Promotor entweder das Sense- ODER das Antisense-Transkript, abhängig von seiner Ausrichtung auf Ihr Gen. |
| T7lac | Hohe Genexpression | Promotor von T7-Bakteriophagen plus Lac-Operatoren | Vernachlässigbare basale Expression, wenn sie nicht induziert wird. Benötigt T7-RNA-Polymerase, die ebenfalls vom lac-Operator gesteuert wird. Kann durch IPTG induziert werden. | Häufig in PET-Vektoren gefunden. Sehr streng von den lac-Betreibern reguliert. Gut zur Modulation der Genexpression durch unterschiedliche Induktorkonzentrationen. |
| Sp6 | in vitro Transkription/ allgemeine Expression | Promotor aus Sp6 Bakteriophagen | Konstitutiv, erfordert aber SP6 RNA Polymerase. | SP6-Polymerase hat eine hohe Prozessivität. Bei Verwendung für die In-vitro-Transkription steuert der Promotor entweder das Sense- ODER das Antisense-Transkript, abhängig von seiner Ausrichtung auf Ihr Gen. |
| araBAD | Allgemeine Expression | Promotor des Arabinose-Stoffwechseloperons | Induzierbar durch Arabinose und unterdrückte Katabolit-Repression in Gegenwart von Glucose oder durch kompetitive Bindung des Antiinduktors Fucose | Schwächer. Häufig in pBAD-Vektoren gefunden. Gut für eine schnelle Regulation und niedrige basale Expression; jedoch nicht gut geeignet für die Modulation der Genexpression durch unterschiedliche Induktorkonzentrationen. |
| trp | Hohe Genexpression | Promotor von E. coli Tryptophan operon | Repressible | Wird mit hohen Konzentrationen von zellulärem Tryptophan ausgeschaltet. |
| lac | Allgemeiner Ausdruck | Promotor von lac operon | Konstitutiv in Abwesenheit von lac Repressor (lacI oder lacIq). Kann durch IPTG oder Lactose induziert werden. | Undichter Promotor mit etwas schwacher Expression. Die lacIq-Mutation erhöht die Expression des Repressors 10x, wodurch die Regulation des Lac-Promotors verschärft wird. Gut zur Modulation der Genexpression durch unterschiedliche Induktorkonzentrationen. |
| Ptac | Allgemeine Expression | Hybridpromotor von lac und trp | Reguliert wie der Lac-Promotor | Enthält -35 Region von trpB und -10 Region von lac. Sehr strenge Regulierung. Gut zur Modulation der Genexpression durch unterschiedliche Induktorkonzentrationen. Im Allgemeinen besserer Ausdruck als lac allein. |
| pL | Hohe Genexpression | Promotor aus Bakteriophagen lambda | Kann temperaturregulierbar sein | Oft gepaart mit dem temperaturempfindlichen cI857-Repressor. |
| T3 | In-vitro-Transkription / allgemeine Expression | Promotor aus T3-Bakteriophagen | Konstitutiv, erfordert jedoch T3-RNA-Polymerase | Bei Verwendung für die In-vitro-Transkription steuert der Promotor entweder das Sense- ODER das Antisense-Transkript, abhängig von seiner Ausrichtung auf Ihr Gen |
Obwohl diese Liste ein guter Anfang ist, vertiefen sich die obigen Tabellen nicht in die gewebe- oder entwicklungsspezifischen Promotoren, die Wissenschaftlern zur Verfügung stehen. Plasmide werden oft therapeutisch eingesetzt, und in diesen Fällen ist es wichtig, die richtigen gewebespezifischen Promotoren zu identifizieren, wie von Forschern des NIH hier beschrieben.
Anmerkung: A. Max Juchheim hat zum Verfassen dieses Artikels beigetragen.