WERBUNG:
Im Kreislaufsystem der Wirbeltiere gibt es zwei Systeme aufwendig verzweigter Röhren, die sich im ganzen Körper verzweigen und Flüssigkeiten zu den Geweben transportieren. Sie sind ein Blutgefäßsystem und ein Lymphsystem. Wenn der Begriff Kreislaufsystem verwendet wird, bezieht er sich nur auf das Blutgefäßsystem.
Das Blutgefäßsystem ist bei Wirbeltieren ein geschlossenes System; Es hat ein kontraktiles Herz und durchgehende Röhren, die Gefäße genannt werden. Das Lymphsystem ist ein offenes System mit Lymphräumen. Die Blutgefäße, die Blut vom Herzen wegführen, sind Arterien, die sich in dünnere Arteriolen teilen und sich in extrem dünne und kleine Kapillaren verzweigen.
Die Wand einer Kapillare besteht aus einer einzigen Schicht quastenförmiger Endothelzellen. Jedes Blutgefäß, einschließlich des Herzens, hat eine Auskleidung von Endothelzellen oder Endothel. Blut kommt nicht direkt mit Gewebezellen in Kontakt.
ANZEIGEN:
Substanzen gelangen von und in Kapillaren durch die Gewebeflüssigkeit, die in Gewebsräumen zwischen Zellen enthalten ist. Der Austausch von Substanzen zwischen den Blut- und Gewebezellen des Körpers erfolgt durch die Kapillarwände. Dies wird durch Pinozytose in den Endothelzellen der Kapillaren bewirkt.
Unter dem Elektronenmikroskop zeigen die Endothelzellen viele kleine Bläschen, die Invaginationen der Plasmamembranen sind. Diese Vesikel bewegen sich von einer Seite der Zelle und geben ihren Inhalt auf der gegenüberliegenden Seite frei.
Somit gibt das Blut Sauerstoff, Nährstoffe und Hormone an die Zellen für den Stoffwechsel ab, und die Zellen geben Wasser, Kohlendioxid und stickstoffhaltige Abfälle an das Blut ab, um sie zu den Ausscheidungsorganen zu transportieren, um sie schnell zu beseitigen.
Die Kapillaren bilden ein Netzwerk in allen Körpergeweben außer Knorpel und Epithel. Von den Kapillaren gelangt das Blut in dünne Venolen, die sich zu Venen verbinden, die Blut zum Herzen transportieren. Aber einige Venen (Pfortader, Nierenvenen und Lebervene) haben Kapillaren, die denen von Arterien ähneln. Aber alles Blut geht nicht durch Kapillaren in Venolen.
ANZEIGEN:
Es gibt auch einige Durchgangskanäle zwischen Arteriolen und Venolen, die in einigen Organen wie der Haut liegen. Es gibt auch arteriovenöse Anastomosen zwischen Arteriolen und Venolen in den Ziffern. Die Funktion solcher Verbindungen ist nicht eindeutig bekannt, obwohl behauptet wird, dass sie Blutdruck und Kreislauf regulieren.
An einigen Stellen findet der Austausch von Material zwischen Geweben und Blut durch dünnwandige Räume oder Sinuskurven statt. In bestimmten Organen kann ein Blutgefäß ein gewundenes Netzwerk winziger Blutgefäße bilden, die als Rete mirabile (Nieren, Luftblase) bezeichnet werden.
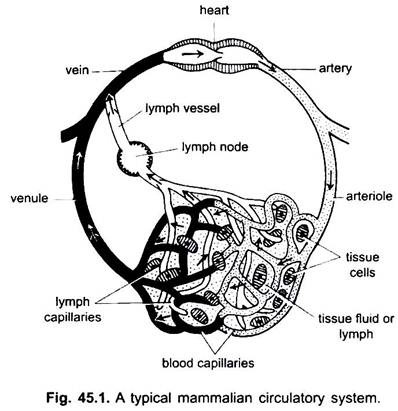
Teile des Kreislaufsystems:
WERBUNG:
Bei Akkordaten ist das Kreislaufsystem geschlossen. Dieser Typ kommt auch in Anneliden (Wirbellosen) vor. Ein anderer Typ ist vom offenen Typ, in dem Kapillaren nicht gefunden werden. Es kommt in Weichtieren und Arthropoden vor. Hier fließt das Blut durch Arterien in verschiedene Organe, die durch Bluträume oder Nebenhöhlen und dann wieder in Gefäße (Venen) zum Herzen gelangen.
Das Kreislaufsystem umfasst Herz, Arterien, Venen, Kapillaren und Blut. Das Herz ist ein modifiziertes Blutgefäß mit Muskelwänden, das sich periodisch zusammenzieht, um das Blut durch bestimmte Gefäße in die verschiedenen Körperteile zu pumpen.
Arterien und ihre Äste bilden ein arterielles System, das das Blut aus dem Herzen transportiert. Venen und ihre Nebenflüsse bilden ein Venensystem, das das Blut aus den Kapillaren von Arterien oder Arteriolen entnimmt und zum Herzen transportiert.
Portalsystem:
Im Portalsystem wird das Blut nicht direkt zum Herzen zurückgeführt, sondern es befindet sich ein zwischengeschaltetes Organ (Leber oder Niere) im Verlauf des zurückkehrenden Blutes. Die Vene, die Blut bringt, beginnt in Kapillaren und endet in Kapillaren, wobei die betreffende Vene sowohl als afferentes als auch als efferentes Gefäß wirkt, die afferenten Gefäße enden wie Arterien in Kapillaren, dann wird das Blut in systemischen Venen gesammelt.
Alle Wirbeltiere haben ein Leberportalsystem, in dem Blut in zwei Sätze von Kapillaren in der Leber gelangt. Untere Wirbeltiere und Embryonen höherer Wirbeltiere haben ebenfalls ein Nierenportalsystem, in dem Blut durch zwei Kapillaren in der Niere fließt, bevor es das Herz erreicht. Die Kapillaren in der Hypophyse bilden ein Hypophysenportalsystem, das ein kleines, aber wichtiges System ist.
Lymphsystem:
Es kommt in Akkordaten außer Cyclostomen und Knorpelfischen vor. Es umfasst Lymphe und Lymphgefäße. Lymphe ist eine Gewebeflüssigkeit, die in den Körperzellen vorkommt. Es ist Blutplasma minus rote Blutkörperchen und einige Proteine.
Lymphkapillaren bilden ein Netzwerk von dünnen Blindendgefäßen, die Lymphe sammeln. Lymphgefäße sind dünnwandige Gefäße, die durch die Vereinigung von Blutkapillaren gebildet werden. Diese leeren sich in Venen. Lymphknoten finden sich auf Lymphgefäßen bei Säugetieren. Diese bilden Lymphozyten aus Blut, die zur Abwehr von Krankheiten eingesetzt werden.
Evolution des Herzens bei Wirbeltieren:
WERBUNG:
Das Herz ist ein ungepaartes Organ, aber sein Ursprung ist bilateral. In einem Embryo bildet das Mesenchym eine Gruppe von Endokardzellen unterhalb des Pharynx. Diese Zellen werden angeordnet, um ein Paar dünner Endothelröhren zu bilden. Die beiden Endothelröhren verschmelzen bald zu einem einzigen Endokardröhrchen, das in Längsrichtung unterhalb des Pharynx liegt.
Das unterhalb des Endoderms liegende splanchnische Mesoderm wird längs um den Endokardschlauch gefaltet. Diese zweischichtige Röhre bildet das Herz, in dem sich das splanchnische Mesoderm verdickt, um ein Myokard oder eine Muskelwand des Herzens und ein äußeres dünnes Epikard oder viszerales Perikard zu bilden. Der Endokardschlauch wird zur Auskleidung des Herzens, das als Endokard bekannt ist.
Falten des splanchnischen Mesoderms treffen sich oben zu einem dorsalen Mesokard, das das Herz im Coelom aufhängt. Bald bildet sich hinter dem Herzen ein transversales Septum, das das Coelom in zwei Kammern unterteilt, eine vordere Perikardhöhle, die das Herz umschließt, und eine hintere Bauchhöhle.
Das Herz ist eine gerade Röhre, aber es nimmt an Länge zu und wird S-förmig, weil seine Enden fixiert sind. Das Auftreten von Klappen, Verengung, Trennwänden im Herzen und differentielle Verdickungen seiner Wände bilden drei oder vier Kammern im Herzen.
1. Einkammerherz:
In Amphioxus (primitives Akkordat) wird kein wahres Herz gefunden. Ein Teil der ventralen Aorta unterhalb des Pharynx ist muskulös und kontraktil und wirkt als Herz.
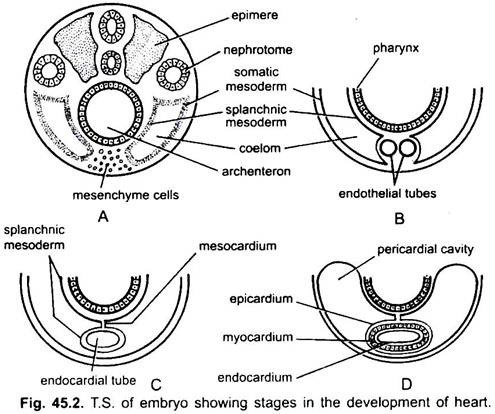
2. Zweikammerherz:
In Cyclostomen sind vier Kammern in linearer Reihenfolge angeordnet – ein dünnwandiger Sinus venosus, ein leicht muskulöser Vorhof (Ohrmuschel), ein muskulöser Ventrikel und ein muskulöser Conus arteriosus oder Bulbus cordis. Es liegt in der Körperhöhle, in der auch andere viszerale Organe vorhanden sind.
Von vier Kammern entsprechen nur Atrium und Ventrikel den vier Kammern (gepaarte Vorhöfe und gepaarte Ventrikel) der höheren Wirbeltiere. In der Evolution des Herzens haben viele Veränderungen stattgefunden.
Elasmobranchen:
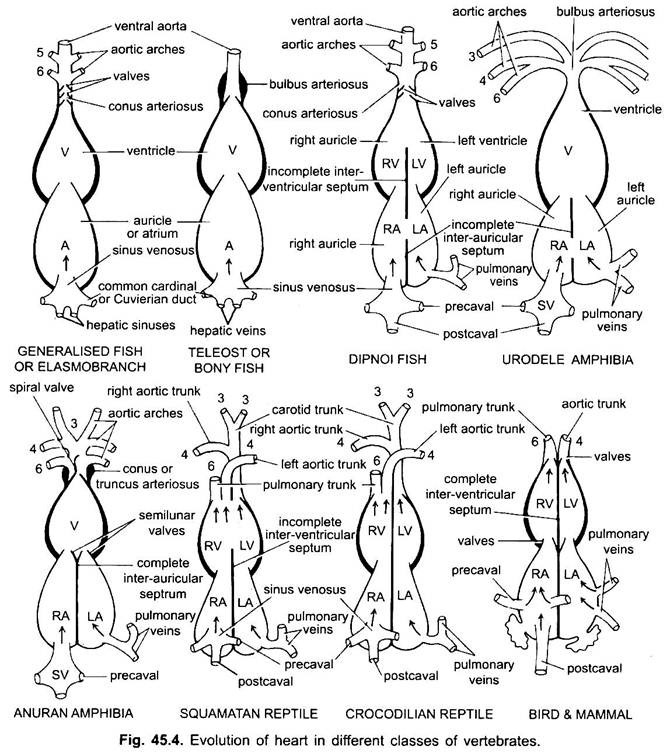
Außer Dipnoi, dem Kreislaufsystem bei Fischen von Cyclostomen bis Teleosts, gelangt nur sauerstofffreies Blut zum Herzen, von dort wird es zu den Kiemen gepumpt, belüftet und dann im Körper verteilt. Das Herz des knorpeligen Dornhai ist muskulös und dorsoventral gebogen S-förmige Röhre mit vier Kompartimenten in einer linearen Reihe.
Sie sind Sinus venosus und Atrium zur Aufnahme von venösem Blut und ein Ventrikel und Conus arteriosus zum Pumpen dieses Blutes. Das Herz ist ein verzweigtes venöses Herz. Der Sinus venosus und der Conus arteriosus sind akzessorische Kammern. Atrium und Ventrikel sind echte Kammern, also ist es ein 2-Kammer-Herz.
Der Sinus venosus öffnet sich anterior in Atrium durch sinu-atriale Öffnung durch ein Paar von Ventilen bewacht. Atrium liegt dorsal zum Ventrikel und öffnet sich ventral in Ventrikel durch eine atrioventrikuläre Öffnung, die von einem Paar von Ventilen bewacht wird. Der dickwandige, muskulöse Ventrikel mündet in einen schmalen Conus arteriosus, der Klappen in zwei Reihen enthält.
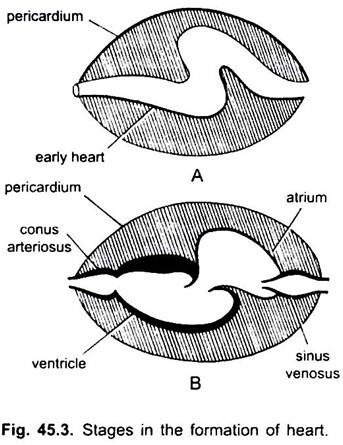
Das Herz ist in der Perikardhöhle eingeschlossen, die durch ein transversales Septum von der Körperhöhle getrennt ist. Conus durchbohrt das Perikard und wird kontinuierlich mit der ventralen Aorta. Die Perikardhöhle kommuniziert mit der Körperhöhle durch zwei Perforationen im Querseptum.
Teleosts:
Ihr Herz ähnelt dem von Clasmobranchen. In Teleosts ist der Konus reduziert und hat ein einzelnes Ventilpaar. Der proximale Teil der ventralen Aorta in der Nähe des Konus wird stark vergrößert und dickwandig, Bulbus arteriosus genannt. Es ist elastisch und dehnt sich zum Zeitpunkt der ventrikulären Kontraktion aus. Das Herz ist also 2-kammerig mit einer einzigen Blutzirkulation.
3. Dreikammerherz:
Dipnoi:
Bei Diphoen teilt ein Septum das Atrium in eine rechte und linke Kammer. Dies korreliert mit der Verwendung der Schwimmblase als Atmungsorgan und stellt den ersten Schritt zur Entwicklung des Kreislaufsystems vom doppelten Typ dar, bei dem sowohl sauerstoffhaltiges als auch nicht sauerstoffhaltiges Blut in das Herz gelangen und getrennt gehalten werden.
Blut aus der rechten Ohrmuschel des Lungenfisches gelangt in den rechten Ventrikel und wird dann durch Lungenarterien, die vom sechsten Paar Aortenbögen abzweigen, in die primitive lungenartige Gasblase gepumpt. Das mit Sauerstoff angereicherte Blut kehrt wie Amphibien über Lungenvenen in den linken Vorhof zurück.
Amphibien:
Bei Amphibien verschiebt sich das dorsale Atrium vor dem Ventrikel. Der Sinus venosus mündet dorsal und nicht posterior in den rechten Vorhof. Das Atrium ist vollständig in rechte und linke Kammern unterteilt und hat kein Foramen ovale im interaurikulären Septum, das in Dipnoen offen bleibt.
In der Ventrikelhöhle entwickeln sich tiefe Taschen. Der Conus arteriosus teilt sich durch eine Spiralklappe in systemische und pulmonale Gefäße. Bei lungenlosen Salamandern ist das interatriale Septum unvollständig und die Lungenvenen fehlen.
Reptilien:
Bei Reptilien ist das Herz weiter fortgeschritten. Das Atrium ist immer vollständig in eine rechte und linke Kammer getrennt, und in vielen Formen ist der Sinus venosus in die Wand des rechten Atriums eingearbeitet. Der Ventrikel ist bei den meisten Reptilien auch teilweise durch ein Septum geteilt, und bei den Alligatoren und Krokodilen ist er vollständig zweikammerig.
Dies bedeutet, dass sauerstoffhaltiges Blut, das von der Lunge zur linken Seite des Herzens kommt, im Wesentlichen von dem nicht sauerstoffhaltigen Blut aus dem Körper zur rechten Seite getrennt ist. So sind bei Krokodilen die beiden Blutarten vollständig getrennt und bei anderen Reptilien fast vollständig, aber in anderen Teilen des Kreislaufsystems kommt es zu einer gewissen Vermischung.
Der embryonale Konus arteriosus teilt sich in drei statt in zwei Gefäße:
(i) Lungenbogen, der Blut von der rechten Seite des Ventrikels in die Lunge transportiert.
(ii) Rechte systemische Aorta, die Blut von der linken Seite des Ventrikels über den rechten vierten Aortenbogen zum Körper transportiert.
(iii) Das linke Herz kommt vom rechten Ventrikel zum linken vierten Aortenbogen.
An der Kontaktstelle mit der systemischen Aorta vom linken Ventrikel aus ist selbst bei Krokodilen eine Öffnung zwischen den beiden vorhanden, die als Foramen von Panizzae bezeichnet wird und in der eine gewisse Vermischung der beiden Blutarten auftreten kann. Somit stellt das Reptilienherz das Übergangsherz gegen das Amphibienherz dar – 2 vollständige Ohrmuscheln und 2 unvollständige Ventrikel mit einer kleinen Vermischung von Blut im rechten und linken Ventrikel.
4. Vierkammerherz:
Aves und Säugetiere:
Bei Vögeln ist der Ventrikel vollständig in zwei Teile geteilt, so dass das Herz vier Kammern hat (2 Ohrmuscheln und 2 Ventrikel). Es gibt eine vollständige Trennung von venösem und arteriellem Blut. Die systemische Aorta verlässt den linken Ventrikel und transportiert Blut zu Kopf und Körper. Während die Lungenarterie den rechten Ventrikel verlässt und Blut zur Sauerstoffversorgung in die Lunge transportiert.
Somit gibt es eine doppelte Zirkulation, in der an keiner Stelle Blut vermischt wird. Der Sinus venosus ist vollständig in die rechte Ohrmuschel eingearbeitet, die zwei präkavale und eine postkavale erhält. Die linke Ohrmuschel erhält sauerstoffreiches Blut durch die Lungenvenen, der Konus arteriosus fehlt, die Lungenaorta entsteht aus dem rechten Ventrikel und die einzelne systemische Aorta entsteht aus dem linken Ventrikel, und beide haben Klappen an ihrer Basis.
Modifikationen der Aortenbögen bei Wirbeltieren:
Embryonale Arterien:
Wenn das Herz in einem Wirbeltierembryo gebildet wird, erscheint ein Blutgefäß namens ventrale Aorta Mitte ventral unter dem Pharynx, der bald mit dem Conus arteriosus verbunden wird. Die vom Herzen ausgehende ventrale Aorta verläuft unter dem Pharynx nach vorne und teilt sich anterior in ein Paar äußerer Halsschlagadern in den Kopf.
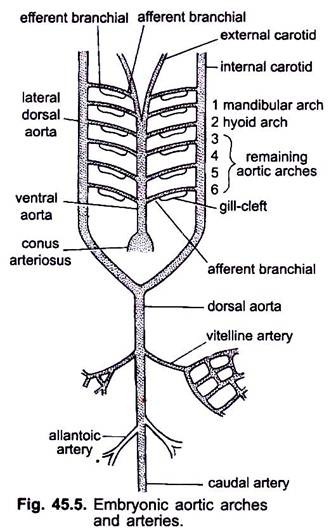
Die ventrale Aorta gibt in gleichem Abstand 6 Paare lateraler Aortenbögen ab, die durch die viszeralen Bögen verlaufen. Jeder Aortenbogen hat eine ventrale afferente Verzweigungsarterie, die venöses Blut zur Kieme trägt, und eine dorsale efferente Verzweigungsarterie, die sauerstoffhaltiges Blut aus der Kieme entnimmt. Die efferenten Verzweigungsarterien beider Seiten verbinden sich dorsal mit der lateralen dorsalen Aorta oder Radix Aorta, die als A. carotis interna in den Kopf eintritt.
Der erste Aortenbogen ist ein Unterkiefer-Aortenbogen, der zweite ist ein Zungenaortenbogen, die übrigen werden als dritter, vierter, fünfter und sechster Aortenbogen bezeichnet. Die laterale dorsale Aorta verschmilzt hinter dem Pharynx zu einer dorsalen Aorta, die sich als Schwanzarterie in der Mitte dorsal in den Schwanz fortsetzt.
Aus der dorsalen Aorta entstehen gepaarte und ungepaarte Arterien, die verschiedene Organe des Körpers versorgen. Bei einem Embryo mit Dottersack entsteht ein Paar vitelliner Arterien aus der dorsalen Aorta und versorgt den Dottersack. Bei Embryonen von Amnioten entsteht ein Paar Nabel- oder Allantoarterien aus der dorsalen Aorta, die das Allantois mit Blut versorgt.
Bei einem Erwachsenen verschmelzen die Vitellinarterien zur Hauptmesenterialarterie, der größte Teil der allantoischen Arterien geht verloren, aber ihre Überreste bilden hypogastrische oder innere Iliakalarterien.
Aortenbögen bei Wirbeltieren:
Bei verschiedenen erwachsenen Wirbeltieren scheint das arterielle System unterschiedlich zu sein, aber sie basieren auf demselben grundlegenden Grundplan. Der Unterschied ist auf die zunehmende Komplexität des Herzens aufgrund einer Veränderung der Atmung von Kiemen zu Lungen zurückzuführen. Die Anzahl der Aortenbögen in der Wirbeltierreihe nimmt progressiv ab.
Cyclostomata:
In Petromyzon gibt es 7 Paare von Aortenbögen. In anderen Cyclostomen variieren diese von 6 Paaren in Myxin und 15 Paaren in Eptatretus.
Fische:
Obwohl sechs als die Grundzahl der Aortenbögen für Fische angesehen wird. Diese Zahl ist sogar bei Haien und Rochen mit dem Verlust des ersten Paares, des Unterkiefer-Aortenbogens, auf fünf reduziert oder wird durch eine efferente Pseudobranchialarterie dargestellt. Bei den meisten Knochenfischen verschwinden sowohl die Aortenbögen des Unterkiefers (i) als auch des Zungenbeins (ii) oder sind stark reduziert.
Bei Polypterus und Dipnoi (Lungenfischen)sind die Kiemen nicht gut entwickelt. Die Lungenarterie entsteht also aus dem efferenten Teil des 6. Bogens auf beiden Seiten, der die Luftblase oder Lunge mit Blut versorgt. In Elasmobranchen und Dipnoi hat jeder Bogen eine afferente und zwei efferente Verzweigungsarterien (gebildet durch Spaltung) in jeder Kieme. Bei Knochenfischen hat jede Kieme eine afferente und eine efferente Arterie.
Bei Tetrapoden brechen Aortenbögen nicht in afferente und efferente Teile auf, da echte innere Kiemen fehlen. Bei allen Tetrapoden verschwinden der erste und der zweite Bogen.
Amphibia:
Hier zeigen die Aortenbögen Veränderungen aufgrund des Verlustes der Kiemen und des Aussehens der Lunge. In Urodelen gibt es äußere Kiemen, die zusätzlich zu den Lungen als Atmungsorgane vorhanden sind. Die Aortenbögen III, IV, V und VI sind vorhanden, obwohl das fünfte Paar in Siren, Amphiuma und Necturus stark reduziert ist. Die Aortenbögen sind in den äußeren Kiemen nicht in afferente und efferente Abschnitte unterteilt, da Äste, die aus IV-, V- und VI-Aortenbögen entstehen, Kapillaren in den äußeren Kiemen bilden.
Die lateralen dorsalen Aorten zwischen den Aortenbögen III und IV bestehen als Gefäßverbindung, dem Ductus caroticus, fort. Der VI-Aortenbogen bildet auf beiden Seiten den pulmo-kutanen Bogen oder die Arterie, die Blut zur Lunge und zur Haut führt. Es behält auch eine Verbindung mit der lateralen dorsalen Aorta, die als Ductus arteriosus (Botalli-Kanal) bekannt ist.
In der Larve von Anuran (Froschkaulquappe) sind die Anordnung der Aortenbögen wie eine erwachsene Urodele aufgrund der Anwesenheit von Kiemen. Bei der Metamorphose verschwinden mit dem Verlust der Kiemen die Aortenbögen I, II und V vollständig, nur die Aortenbögen IIIrd, IV und Vlth sind vorhanden. Die laterale dorsale Aorta zwischen dem dritten und vierten Aortenbogen (Ductus caroticus) verschwindet ebenfalls. Somit wird der dritte Aortenbogen zusammen mit einem Teil der ventralen Aorta zum Carotis-Bogen, der sauerstoffreiches Blut in die Kopfregion trägt.
Der vierte Aortenbogen entlang seiner lateralen dorsalen Aorta bildet den Systembogen. Der sechste Aortenbogen wird zum pulmokutanen Bogen, der Lunge und Haut mit venösem Blut versorgt. Der Ductus arteriosus verschwindet während der Metamorphose. So haben erwachsene Anurane nur III, IV und VI Aortenbögen. Diese werden auch von Amnioten zurückgehalten.
Reptilien:
Bei Reptilien werden die Kiemen vollständig durch Lungen ersetzt. Nur III, IV und VI Aortenbögen sind vorhanden. Mit der teilweisen Trennung des Ventrikels in zwei Teile werden der distale Teil des Conus arteriosus und die gesamte ventrale Aorta in drei Gefäße aufgeteilt, d. H. Zwei Aorten- oder systemische und eine pulmonale. Der rechte Systembogen (IV) entsteht aus dem linken Ventrikel, der sauerstoffreiches Blut zum Karotisbogen (III) transportiert. Die linken systemischen (IV) und pulmonalen Aorten (VI) haben ihren Ursprung im rechten Ventrikel. Die linke Arterie transportiert desoxygeniertes oder gemischtes Blut durch die dorsale Aorta zum Körper.
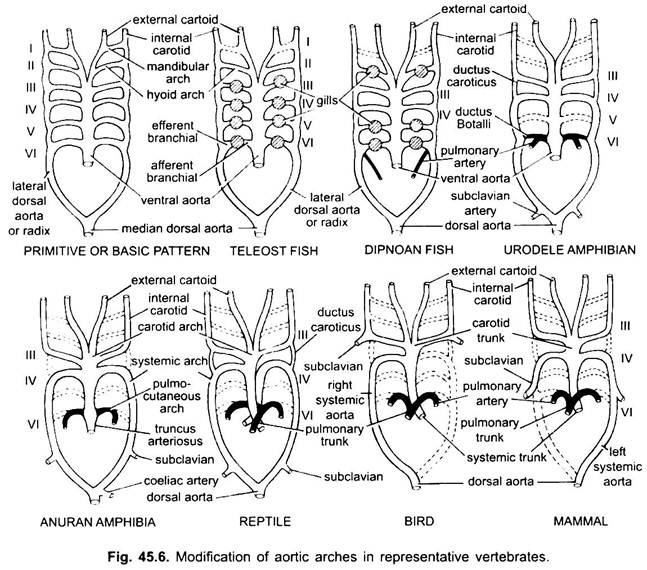
Während die Lungenarterie sauerstoffarmes Blut in die Lunge transportiert. Der Ductus caroticus verschwindet, bleibt aber bei Schlangen und einigen Eidechsen (Uromastix) bestehen. Der Ductus arteriosus verschwindet bei den meisten Reptilien, obwohl er bei Sphenodon und einigen Schildkröten in reduzierter Form erhalten bleibt. Aufgrund der Vermischung von Blut sind Reptilien kaltblütige Tiere wie Fische und Amphibien.
Aves:
Bei Vögeln sind die Aortenbögen III, IV und VI vorhanden. Sie folgen dem allgemeinen Muster der Reptilien mit einigen Unterschieden. Mit der vollständigen Teilung des Ventrikels in zwei Teile haben sich der Conus arteriosus und die ventrale Aorta zu zwei Gefäßen geteilt, einer systemischen Aorta aus dem linken Ventrikel und einer pulmonalen Aorta aus dem rechten Ventrikel.
Der dritte Aortenbogen mit Resten der lateralen und ventralen Aorten bildet die Karotis, die aus der systemischen Aorta entsteht. Der vierte Aortenbogen bildet die systemische Aorta nur auf der rechten Seite. Es verbindet sich mit der lateralen Aorta seiner eigenen Seite und bildet die dorsale Aorta. Ein Teil des vierten Aortenbogens der linken Seite bildet die linke A. subclavia, der Rest verschwindet zusammen mit seiner lateralen dorsalen Aorta. Der sechste Aortenbogen bildet die Lungenaorta. Ductus caroticus und Ductus arteriosus verschwinden.
Säugetiere:
Bei Säugetieren bestehen auch die III, IV und VI aoric Bögen fort. Der Ventrikel ist vollständig in zwei Teile geteilt.
Der Conus arteriosus und die ventrale Aorta teilen sich zu zwei Gefäßen:
(i) Eine systemische Aorta aus dem linken Ventrikel und
(ii) Eine pulmonale Aorta aus dem rechten Ventrikel. Dritter Aortenbogen mit Resten der lateralen und ventralen Aorten bildet den Carotis-Bogen.
Der vierte Aortenbogen bildet nur auf der linken Seite die systemische Aorta, während auf der rechten Seite sein proximaler Teil eine innominate und rechte Arteria subclavia bildet, der Rest zusammen mit seiner lateralen dorsalen Aorta verschwindet. Sechster Aortenbogen bildet die Lungenaorta. Der Ductus arteriosus degeneriert, bleibt aber bei einigen bis zum Schlüpfen oder zur Geburt in reduzierter Form auf der linken Seite als dünnes Ligamentum arteriosum bestehen.
Venensystem:
Embryonale Venen:
Bei allen Wirbeltierembryonen ist das Venensystem einfach und ähnlich. Die Venen sind meist gepaart und symmetrisch angeordnet. Bei Embryonen ohne Dottersack bildet sich im splanchnischen Mesoderm unterhalb des Darms eine subintestinale Vene.
Es schlingt um den Anus und wird posterior fortgesetzt als:
(i) Eine Schwanzvene in den Schwanz. In allen Embryonen mit einem Dottersack (ob mit Eigelb oder nicht) entsteht ein Paar vitelliner Venen aus dem Dottersack und verbindet sich mit dem hinteren Teil des sich entwickelnden Herzens, der zum Sinus venosus wird. Tatsächlich ist die Verschmelzung dieser Vitalvenen für die Herzbildung bei Knochenfischen, Reptilien und Vögeln verantwortlich. Jede vitelline Vene an ihrem hinteren Ende verbindet sich mit der subintestinalen Vene, die auf die gleiche Weise gebildet wird wie bei den Embryonen ohne Dottersack.
(ii) Zwischen den Nieren entsteht ein Paar subkardinaler Venen, die sich mit der Schwanzvene verbinden.
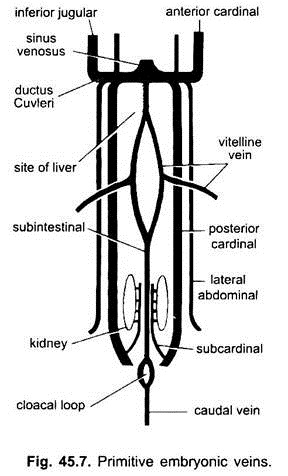
( iii) Es bilden sich gepaarte vordere und hintere Kardinalvenen, die das Blut vom Kopf bzw. vom hinteren Teil des Körpers tragen. Die vorderen und hinteren Kardinalvenen jeder Seite vereinigen sich zu einem Ductus Cuvieri oder einer gemeinsamen Kardinalvene, die durch das Querseptum nach innen verläuft, um in den Sinus venosus einzutreten.
Bei Fischen und Salamandern (Urodeles)kommt eine Vena jugularis inferior von der ventralen Seite des Kopfes, um sich der Vena cardinalis communis anzuschließen. Es hat kein Homolog in anderen Wirbeltieren. Bei einem Amnioten kommt ein Paar lateraler oder ventraler Bauchvenen von der Körperwand, um in die gemeinsamen Kardinalvenen einzutreten.
Fische:
Gemeinsame Kardinalvene (Ductus Cuvier) tritt von jeder Seite in den Sinus venosus ein und wird durch die Fusion von vorderen und hinteren Kardinälen gebildet. Blut aus dem Kopf wird von den vorderen Kardinälen gesammelt, und Blut aus den Nieren und Gonaden wird von den hinteren Kardinälen gesammelt.
In die Cuvier-Gänge gelangen auch die gepaarten lateralen Bauchvenen, die Blut von der Körperwand und den gepaarten Anhängseln aufnehmen. Das Nierenportalsystem besteht aus der Schwanzvene und zwei seitlich der Nieren gelegenen Nierenportalvenen, die in den Nieren kapillarisierten.
Das Leberportalsystem transportiert Blut aus Magen und Darm und gibt es in die Leber zurück, wo es nach Durchlaufen einer Reihe von Sinusoiden über gepaarte rheumatische Venen in den Sinus venosus gelangt.
Bei Teleosts fehlen die lateralen Bauchvenen, so dass Blut aus den Subclavians, das die Brustanhänge entwässert, direkt in den Sinus venosus gelangt und das Blut aus den Beckenvenen, das die Beckenanhänge entwässert, in die Postkardinale gelangt.
Bei Dipnoen ist eine einzelne ventrale Bauchvene vorhanden, die vermutlich aus einer Fusion der lateralen Bauchmuskeln stammt. Diese Vene erhält Blut von den Ilacs über gepaarte Beckenvenen und tritt in den rechten Kanal von Cuvier ein. Darüber hinaus tritt aus dem rechten postkardinalen System eine neue Vene, die Postclaval, auf, die bei höheren Wirbeltieren von großer Bedeutung ist. Es ist mit der Schwanzvene verbunden und gelangt durch die Leber zum Sinus.
Bei Embryonen von Amnioten sind die seitlichen Bauchvenen als Nabelvenen oder allantoische Venen bekannt, da sie die Allantois entwässern. Die Vitellin- und Nabelvenen gehen bei der Geburt verloren, nur ihre Überreste bleiben bestehen.
Modifikationen der Venen:
Wie bei Arterien sind die Venen verschiedener Wirbeltiere auf demselben Grundriss angeordnet. Alle von ihnen gezeigten Variationen folgen einer Sequenz in der Wirbeltierreihe. In seiner Entwicklung durchläuft das Venensystem höherer Wirbeltiere die Stadien, die in Embryonen niedrigerer Formen zu sehen sind.
1. Wenn die Leber gebildet wird, bildet der proximale Teil der Vitellinvenen oder Subintestinalvenen die Lebervenen zwischen der Leber und dem Herzen, der distale Teil der linken Vitellinvene oder Subintestinalvene wird zur Leberportalvene, die Sinusoide in der Leber bildet, um ein in allen vorhandenes Leberportalsystem entstehen zu lassen.
2. Vordere Kardinalvenen bestehen als innere Jugularvenen fort.
3. Außer bei Fischen werden die gemeinsamen Kardinalvenen zu den präkavalen Venen, die in den Sinus venosus (Amphibien, Reptilien) oder die rechte Ohrmuschel (Vögel, Säugetiere) gelangen. In jedem Vorderbein bildet sich eine Vena subclavia, die sich mit der Vena precaval verbindet.
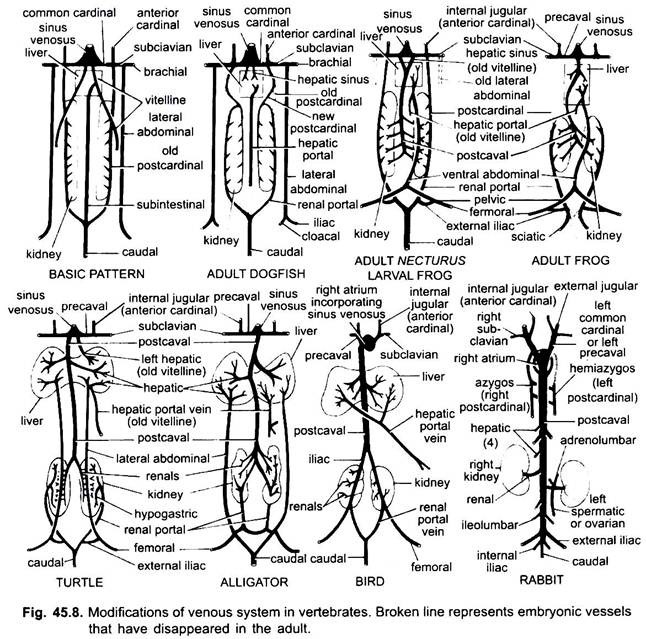
4. Die Vena caudalis verliert ihre Verbindungen mit den Vena subintestinalis und Vena subcardinalis, ihr vorderer Teil teilt sich in zwei Äste auf, die bei allen außer Säugetieren die hinteren Kardinalvenen verbinden.
5. Die hinteren Kardinalvenen bestehen bei Fischen als solche fort, aber bei anderen zerfällt jeder in zwei Teile, ein vorderer Teil, der bei Amphibien, Reptilien und Vögeln verschwindet, aber bei Säugetieren bildet der rechte vordere Teil eine Azygosvene, der linke kann eine Hemiazygosvene bilden, die ihre Verbindung mit der Präkaval verliert und sich durch eine transversale Anastomose mit der Azygosvene verbindet; Die hinteren Teile der hinteren Kardinalvenen, die sich der Schwanzvene angeschlossen haben, werden zu den Nierenportalvenen.
Die Nierenportalvene jeder Seite bildet Kapillaren in der Niere, um ein Nierenportalsystem zu bilden, das bei Fischen, Amphibien und Reptilien vollständig ist, bei Vögeln jedoch stark reduziert wird und bei Säugetieren fehlt, da die hinteren Teile der hinteren Kardinalvenen bei Säugetieren verschwinden.
6. Die Nierenportalvenen von Fischen sammeln Blut nur vom Schwanz, aber bei Amphibien, Reptilien und Vögeln usurpieren sie auch die Venen von den Beinen.
7. Bei einigen Fischen und allen Tetrapoden verbinden sich die vitellinen Venen mit der subkardinalen Vene, um eine postkavale Vene zu bilden. Bei Amphibien und Reptilien erstreckt sich die Vena postcavalis bis zu den hinteren Enden der Nieren, bei Vögeln verbindet sie sich mit den Nierenportalvenen, wodurch das Nierenportalsystem reduziert wird, bei Säugetieren verbindet die Vena postcavalis die Venen von den Beinen und dem Schwanz, so dass das Nierenportalsystem vollständig eliminiert wird.
8. Die beiden seitlichen Abdominalvenen bestehen als solche bei Fischen fort, aber von Dipnoi aufwärts verschmelzen sie zu einer vorderen Abdominalvene, die sich in der Nähe der Leber mit der Leberportalvene verbindet, die vordere Abdominalvene verbindet somit das Nierenportal und das Leberportalsystem. In jedem Hintergliedmaßen bildet sich eine Iliakalvene, die sich mit der vorderen Bauchvene verbindet. Die vordere Bauchvene beginnt bei Reptilien an Bedeutung zu verlieren, bei Vögeln wird sie modifiziert, um die epigastrischen und steißbein-mesenterialen Venen zu bilden, bei Säugetieren verschwindet sie, außer bei Tachyglossus.
9. Bei luftatmenden Formen wachsen Lungenvenen aus der linken Ohrmuschel heraus und gelangen in die Lunge.
10. Die Nabelvenen (allantoischen Venen) des Embryos verschwinden, wenn die Lunge als Atmungsorgane funktionsfähig wird.
Tetrapode:
Das Venensystem von Amphibien ähnelt sehr dem von Lungenfischen, mit der Ausnahme, dass die Bauchvene eher in das Leberportalsystem als in den Sinus venosus eintritt. Die vorderen Kardinäle bestehen bei allen erwachsenen Tetrapoden als innere Jugularvenen fort. Inferiore Jugularvenen fehlen. Gemeinsame Kardinäle werden die vordere Hohlvene oder Präkavalen, die Sinus venosus bei Amphibien und Reptilien verbinden.
Bei Vögeln und Säugetieren dringen Präkavalen direkt in die rechte Herzmuschel ein, da der Sinus venosus fehlt. Bei Mann, Katze usw., linker precaval ist verloren, so dass das Blut der linken Seite durch einen brachio-cephalic Zweig in den rechten precaval eintritt. Bei lungenlosen Salamandern fehlen die Lungenvenen, da das interatriale Septum unvollständig ist.
Die Postkardinalvenen verbinden sich posterior mit der Schwanzvene und anterior mit den Präkavalen, um die gemeinsamen Kardinäle bei Larvenfrosch und Nekturus zu bilden.
Bei Reptilien kommt es zu einer stärkeren Entwicklung der Lungenvenen und der postkavalen Vene und zu einer Verringerung der Bedeutung des Nierenportalsystems, das Blut aus dem hinteren Teil des Körpers in die Nieren bringt.
Bei Vögeln gibt es zwei funktionelle Präkavalvenen, die durch die Vereinigung von Jugularvene und Subclavia auf jeder Seite gebildet werden, und eine vollständige Postkavalvene. Es erhält Blut von den Gliedmaßen über die Nierenportale, die durch die Nieren verlaufen, aber nicht in Kapillaren zerfallen und nicht mit den Nierenportalen der unteren Wirbeltiere vergleichbar sind.
Bei Säugetieren kann es eine oder zwei präkavale Venen geben. Es gibt eine einzige postkavale Vene, deren embryologische Entwicklung sehr kompliziert ist. Alle Hohlvenen treten direkt in die rechte Ohrmuschel ein, da der Sinus venosus im embryonalen Leben in die Wand dieser Herzkammer absorbiert wird. Es gibt kein Nierenportalsystem, obwohl das Leberportalsystem dem anderer Wirbeltiere sehr ähnlich ist.