reclame:
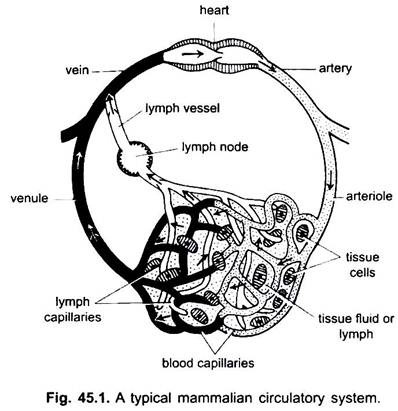
In de bloedsomloop van gewervelde dieren zijn er twee systemen van uitgebreid vertakkende buizen, die ramify door het lichaam en dragen vloeistoffen naar de weefsels. Ze zijn een bloedvatsysteem en een lymfatisch systeem. Wanneer de term bloedsomloop wordt gebruikt, verwijst deze alleen naar het bloedvatstelsel.
het bloedvatstelsel is een gesloten systeem bij gewervelde dieren; het heeft een contractiel hart en continue buisjes die vaten worden genoemd. Het lymfestelsel is een open systeem met lymferuimten. De bloedvaten die het bloed van het hart wegvoeren zijn slagaders, die zich in dunnere arteriolen verdelen, vertakt in extreem dunne en kleine haarvaten.
de wand van een capillair bestaat uit een enkele laag kwastvormige endotheelcellen. Elk bloedvat, inclusief het hart, heeft een bekleding van endotheelcellen of endotheel. Bloed komt niet direct in contact met weefselcellen.
reclame:
stoffen gaan van en in de haarvaten door de weefselvloeistof in weefselruimten tussen cellen. Uitwisseling van stoffen tussen de bloed-en weefselcellen van het lichaam vindt plaats via de capillaire wanden. Dit wordt veroorzaakt door pinocytose in de endothelial cellen van haarvaten.
onder een elektronenmicroscoop vertonen de endotheelcellen vele kleine blaasjes die invaginaties zijn van de plasmamembranen. Deze blaasjes bewegen van één kant van de cel en geven hun inhoud aan de andere kant vrij.
zo geeft het bloed zuurstof, voedingsstoffen en hormonen af aan de cellen voor het metabolisme, en geven de cellen water, kooldioxide en stikstofhoudend afval in het bloed af om deze naar de uitscheidingsorganen te voeren voor een snelle eliminatie.
de haarvaten vormen een netwerk in alle lichaamsweefsels behalve kraakbeen en epitheel. Vanuit de haarvaten gaat het bloed over in dunne venules die samen aderen vormen die bloed naar het hart voeren. Maar sommige aderen (portale aderen, renale aders, en hepatische ader) hebben haarvaten die net als die van slagaders. Maar al het bloed gaat niet door haarvaten in venules.
reclame:
er zijn ook enkele door kanalen tussen arteriolen en venules die in sommige organen, zoals de huid liggen. Er zijn ook arterioveneuze anastomoses tussen arterioles en venules die in de cijfers worden gevonden. De functie van dergelijke verbindingen is niet duidelijk bekend, hoewel wordt beweerd dat ze de bloeddruk en de bloedsomloop reguleren.
op sommige plaatsen vindt de uitwisseling van materiaal tussen weefsels en bloed plaats via dunwandige ruimten of sinusoïden. In bepaalde organen kan een bloedvat een opgerold netwerk van kleine bloedvaten vormen genaamd rete mirabile (nieren, luchtblaas).
Delen van de bloedsomloop:
reclame:
in chordaten is de bloedsomloop van een gesloten type. Dit type komt ook voor bij anneliden (ongewervelde dieren). Een ander type is van open type waarin haarvaten niet worden gevonden. De soort komt voor in weekdieren en geleedpotigen. Hier stroomt het bloed door slagaders naar verschillende organen, die door bloedruimten of sinussen gaan en dan weer in bloedvaten (aderen) naar het hart.
de bloedsomloop omvat het hart, de slagaders, de aderen, de haarvaten en het bloed. Hart is een gemodificeerd bloedvat met Gespierde wanden, die periodiek samentrekt om het bloed in de verschillende delen van het lichaam te pompen via bepaalde bloedvaten. Slagaders en hun vertakkingen vormen een arterieel systeem dat het bloed uit het hart transporteert. Aderen en hun zijrivieren vormen een veneus systeem dat het bloed uit de haarvaten van slagaders of arteriolen neemt en dat naar het hart voert.
portaalsysteem:
in het portaalsysteem wordt het bloed niet rechtstreeks naar het hart teruggestuurd, maar er is een interposerend orgaan (lever of nier) in de loop van het terugkerende bloed. De ader die bloed brengt begint in haarvaten en eindigt in haarvaten, de betrokken ader werkt zowel als afferente en efferente vat, de afferente vaten eindigen in haarvaten net als slagaders, dan wordt het bloed verzameld in systemische aderen.
alle gewervelde dieren hebben een leverportaalsysteem waarbij het bloed in twee reeksen haarvaten in de lever overgaat. De lagere gewervelde dieren en de embryo ‘ s van hogere gewervelde dieren hebben een nierportaalsysteem ook waarin bloed door twee reeksen haarvaten in de nier alvorens het hart te bereiken overgaat. De haarvaten in de hypofyse vormen een slijmachtig portaalsysteem dat een klein maar belangrijk systeem is.
lymfestelsel:
het komt voor in chordaten, met uitzondering van cyclostomen en kraakbeenvissen. Het omvat lymfe en lymfevaten. Lymfe is een weefsel vloeistof gevonden tussen de lichaamscellen. Het is bloedplasma minus rode bloedlichaampjes en sommige eiwitten.
Lymfecapillairen die een netwerk vormen van dunne, blinde bloedvaten, die lymfe verzamelen. Lymfevaten zijn dunwandige vaten die worden gevormd door de Vereniging van bloedcapillairen. Deze legen in aderen. Lymfeklieren worden gevonden op lymfevaten bij zoogdieren. Deze vormen lymfocyten van bloed die worden gebruikt voor de afweer van het lichaam tegen ziekten.
evolutie van het hart bij gewervelde dieren:
reclame:
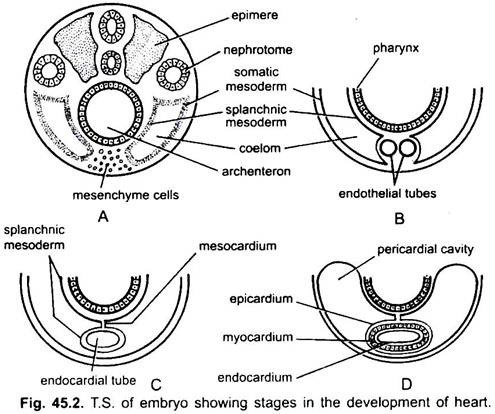
het hart is een ongepaarde orgaan, maar zijn oorsprong is bilateraal. In een embryo vormt het mesenchyme een groep endocardiale cellen onder de keelholte. Deze cellen worden geschikt om een paar dunne endothelial buizen te vormen. De twee endotheliale buizen smelten snel samen tot een enkele endocardiale buis die in de lengterichting onder de keelholte ligt.
het splanchnische mesoderm dat onder de endoderm ligt, wordt in de lengterichting om de endocardiale buis gevouwen. Deze tweelaagse buis zal het hart vormen waarin het splanchnische mesoderm verdikt tot een myocardium of spierwand van het hart en een buitenste dunne epicardium of visceraal pericardium. De endocardiale buis wordt de bekleding van het hart bekend als endocardium.
plooien van splanchnisch mesoderm komen boven samen om een dorsaal mesocardium te vormen dat het hart in de coelom ophangt. Al snel wordt een transversaal septum gevormd achter het hart dat de coelom verdeelt in twee kamers, een voorste pericardiale holte omsluit het hart en een achterste buikholte.
het hart is een rechte buis, maar het neemt in lengte toe en wordt S-vormig omdat de uiteinden vast zijn. Verschijning van kleppen, vernauwing, wanden in het hart, en differentiële verdikkingen van de wanden vormen drie of vier kamers in het hart.
1. Eenkamervormig hart:
in Amphioxus (primitief akkoord) wordt geen echt hart gevonden. Een deel van de ventrale aorta onder de keelholte is gespierd en contractiel en fungeert als hart.
2. Hart Met Twee Kamers:
bij cyclostomen zijn er vier kamers in lineaire volgorde gerangschikt – een dunwandige sinus venosus, een licht Gespierd atrium( oorschelp), een Gespierde ventrikel en een Gespierde conus arteriosus of bulbus cordis. Het ligt in de lichaamsholte waarin ook andere viscerale organen aanwezig zijn. Van de vier kamers komen alleen atrium en ventrikel overeen met de vier kamers (gepaarde atria en gepaarde ventrikels) van de hogere gewervelde dieren. In de evolutie van het hart hebben vele veranderingen plaatsgevonden.
Elasmobranchen:
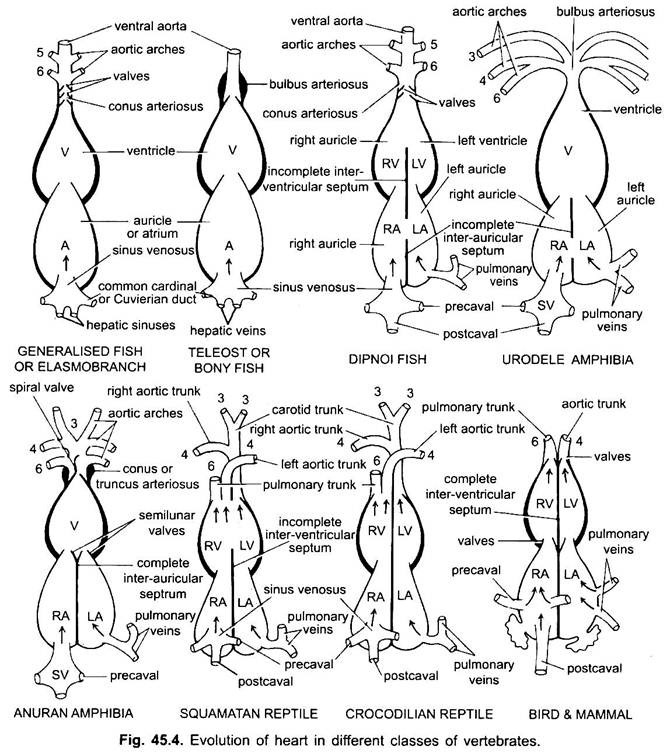
behalve Dipnoi, de bloedsomloop bij vissen van cyclostomen tot teleost, gaat alleen ongeoxygeneerd bloed naar het hart, van daaruit wordt het naar de kieuwen gepompt, belucht en vervolgens naar het lichaam gedistribueerd. Het hart van kraakbeenhaai is gespierd en dorsoventraal gebogen S-vormige buis met vier compartimenten in een lineaire reeks.
het zijn sinus venosus en atrium voor het ontvangen van veneus bloed, en een ventrikel en conus arteriosus voor het pompen van dit bloed. Het hart is een vertakt veneus hart. De sinus venosus en conus arteriosus zijn bijkomende kamers. Atrium en ventrikel zijn echte kamers, dus het is een hart met 2 kamers.
de sinus venosus opent anterioraal in het atrium door een sinuatriale opening die wordt bewaakt door een paar kleppen. Atrium ligt dorsaal naar ventrikel en opent ventraal in ventrikel door een atrioventriculaire opening bewaakt door een paar kleppen. De dikwandige, Gespierde ventrikel opent in een smalle conus arteriosus die kleppen in twee reeksen bevat.
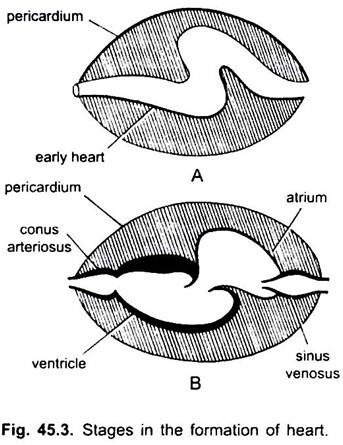
het hart is ingesloten in de pericardiale holte, gescheiden van de lichaamsholte door een dwars septum. Conus doorboort het pericardium en wordt continu met de ventrale aorta. Pericardiale holte communiceert met de lichaamsholte door middel van twee perforaties in het transversale septum.
Teleost:
hun hart lijkt op dat van clasmobranchen. Bij teleosts is de conus gereduceerd en heeft een enkel paar kleppen. Het proximale deel van de ventrale aorta dicht bij conus wordt sterk vergroot en dikwandig, genaamd bulbus arteriosus. Het is elastisch en verwijdt op het moment van ventriculaire contractie. Het hart is dus tweekamervormig met één bloedcirculatie.
3. Hart met drie kamers:
Dipnoi:
bij tweespalt verdeelt een septum het atrium in een rechter-en linkerkamer. Dit is gecorreleerd met het gebruik van de zwem-blaas als een orgaan van ademhaling en vertegenwoordigt de eerste stap naar de ontwikkeling van de dubbel-type bloedsomloop waarbij zowel zuurstofrijk en niet-zuurstofrijk bloed in het hart en gescheiden worden gehouden.
bloed van de rechteroorschelp van de longvis gaat over in de rechterhartkamer en wordt vervolgens in de primitieve longachtige gasblaas gepompt door de longslagaders die zich aftakken van het zesde paar aortabogen. Het zuurstofrijk bloed keert terug naar het linker atrium via longaders zoals amfibieën.
amfibie:
bij amfibieën verschuift het dorsale atrium anterior naar ventrikel. De sinus venosus opent dorsaal in het rechter atrium en niet achteraf. Het atrium is volledig verdeeld in rechter en linker kamers en heeft geen foramen ovale in het Inter-auriculair septum, dat open blijft in dipnoans.
diepe zakken ontwikkelen zich in de ventriculaire holte. De conus arteriosus verdeelt zich in systemische en pulmonale vaten door een spiraalklep. Bij longsalamanders is het interatriale septum onvolledig en zijn de longaders afwezig.
Reptilia:
bij reptielen is het hart verder gevorderd. Het atrium is altijd volledig gescheiden in een rechter en linker kamer, en in vele vormen is de sinus venosus opgenomen in de wand van het rechter atrium. De ventrikel is ook gedeeltelijk gedeeld door een septum in de meeste reptielen, en in de alligators en krokodillen is volledig twee-kamered.
dit betekent dat zuurstofrijk bloed afkomstig van de longen naar de linkerkant van het hart in wezen gescheiden is van het zuurstofvrije bloed van het lichaam naar de rechterkant. Bij krokodilachtigen zijn de twee soorten bloed volledig gescheiden, en bij andere reptielen bijna compleet, maar in andere delen van de bloedsomloop is er wel enige vermenging.
de embryonale conus arteriosus splitst zich in drie in plaats van twee vaten:
(i) pulmonale boog die bloed naar de longen van de rechterkant van het ventrikel voert.
(ii) rechter systemische aorta die bloed van de linkerkant van het ventrikel naar het lichaam voert via de vierde rechter aortaboog.
(iii) links systemisch komt van de rechter ventrikel naar de linker vierde aortaboog.
op het punt van contact met de systemische aorta van de linker hartkamer, zelfs bij krokodilachtigen, is er een opening tussen de twee aanwezig, het foramen van Panizzae genaamd, waar de twee soorten bloed enigszins kunnen worden gemengd. Dus, reptiliaans hart vertegenwoordigt de overgang hart tegen amfibie hart-2 volledige oorschelpen en 2 onvolledige ventrikels met een beetje mengen van bloed in rechts en links systemische.
4. Vierkamerhart:
Aves en Mammalia:
bij vogels is het ventrikel volledig verdeeld in twee kamers, zodat het hart vierkamervormig is (2 auricles en 2 ventrikels). Er is volledige scheiding van veneus en arterieel bloed. De systemische aorta verlaat de linker hartkamer en draagt bloed naar het hoofd en lichaam. Terwijl de longslagader de rechter ventrikel verlaat en bloed naar de longen voert voor oxygenatie.
er is dus een dubbele bloedsomloop waarbij op geen enkele plaats bloed wordt gemengd. De sinus venosus is volledig opgenomen in de rechter oorschelp, die twee precavals en een postcaval ontvangt. De linker oorschelp ontvangt zuurstofrijk bloed door longaderen, conus arteriosus afwezig is, de longaorta ontstaat uit de rechter ventrikel, en één systemische aorta ontstaat uit de linker ventrikel, en beide hebben kleppen op hun basissen.
modificaties van Aortakragen bij gewervelde dieren:
embryonale slagaders:
wanneer het hart wordt gevormd in een embryo van gewervelde dieren, verschijnt een bloedvat genaamd ventrale aorta Midden ventraal onder de keelholte, die al snel met de conus arteriosus verbonden raakt. De ventrale aorta die ontstaat uit het hart loopt naar voren onder de keelholte en verdeelt anteriorly in een paar externe halsslagaders in het hoofd.
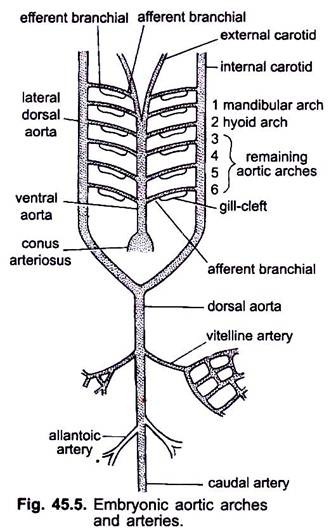
de ventrale aorta geeft op gelijke afstand 6 paar laterale aortabogen af die door de viscerale bogen lopen. Elke aortaboog heeft een ventrale afferente branchiale slagader die veneus bloed naar de kieuw voert en een dorsale efferente branchiale slagader die zuurstofrijk bloed uit de kieuw neemt. De efferent branchial slagaders van beide zijden verbinden dorsaal met de laterale dorsale aorta of radix aorta die in het hoofd als interne halsslagader komt.
de eerste aortaboog is een mandibulaire aortaboog, de tweede is een tongbeen aortaboog, de overige worden derde, vierde, vijfde en zesde aortaboog genoemd. De laterale dossale aorta fuseren achter de keelholte tot een dorsale aorta die wordt voortgezet Midden-dorsaal in de staart als een caudale slagader. Uit de dorsale aorta ontstaan gepaarde en niet-gepaarde slagaders die verschillende organen van het lichaam voeden. In een embryo met een dooierzak ontstaat een paar vitelline slagaders uit de dorsale aorta en levert de dooierzak. In embryo ‘ s van vruchtwater komt een paar navelstreng of allantoic slagaders uit de dorsale aorta voort die bloed aan de allantois leveren.
bij een VOLWASSENE fuseren de vitelline slagaders tot de belangrijkste mesenterische slagaders, het grootste deel van de allantoïsche slagaders gaat verloren, maar de restanten daarvan vormen hypogastrische of interne iliacale slagaders.
Aortabogen bij gewervelde dieren:
bij verschillende volwassen gewervelde dieren lijkt het arteriële systeem verschillend, maar ze zijn gebouwd op hetzelfde fundamentele plan. Het verschil is te wijten aan toenemende complexiteit van het hart als gevolg van een verandering in de ademhaling van kieuwen naar longen. Er is een progressieve vermindering van het aantal aorta bogen in de gewervelde series.
Cyclostomata:
in Petromyzon zijn er 7 paar aortabomen. Bij andere cyclostomen variëren deze van 6 paar in Myxine en 15 paar in Eptatretus.
vissen:
hoewel zes wordt beschouwd als het basisaantal aortabomen voor vissen. Dit aantal wordt teruggebracht tot vijf zelfs bij haaien en roggen met het verlies van het eerste paar, de mandibulaire aortaboog of het wordt vertegenwoordigd door een efferente pseudobranchiale slagader. Bij de meeste benige vissen verdwijnen de onderkaak (i) en het tongbeen (ii) aortabomen of worden ze veel kleiner.
bij Polypterus en Dipnoi (longvissen) zijn kieuwen niet goed ontwikkeld. Dus de longslagader ontstaat uit het efferente deel van de 6e boog aan weerszijden die bloed levert aan de luchtblaas of long. Bij elasmobranchs en Dipnoi heeft elke boog één afferente en twee efferente vertakte slagaders (gevormd door splitsing) in elke kieuw. Bij benige vissen heeft elke kieuw een afferente en een efferente slagader.
bij tetrapoda breken aortakragen niet op in afferente en efferente delen omdat echte inwendige kieuwen ontbreken. In alle tetrapoda verdwijnen eerst en seconde bogen.
amfibie:
hier vertonen de aortabooggen modificatie als gevolg van het verlies van kieuwen en het uiterlijk van de longen. In urodeles zijn er uitwendige kieuwen aanwezig als ademhalingsorganen naast de longen. De III, IV, V en VI aortabomen zijn aanwezig, hoewel het vijfde paar sterk gereduceerd is in Siren, Amphiuma en Necturus. De aorta bogen worden niet gebroken in de uitwendige kieuwen in afferente en efferente porties, omdat takken die voortvloeien uit IV, V en VI aorta bogen vormen haarvaten in de uitwendige kieuwen.
de laterale dorsale aortae tussen de III en IV aorta bogen blijven bestaan als een vasculaire verbinding, de ductus caroticus. De VI aortaboog vormt de pulmo-cutane boog of slagader aan beide zijden die bloed naar de long en de huid. Het behoudt ook een verbinding met de laterale dorsale aorta bekend als een ductus arteriosus (kanaal van Botalli).
in de larve van anuran (kikkervisje) zijn de arrangementen van aortabomen als een volwassen urodele door de aanwezigheid van kieuwen. Bij metamorfose, met het verlies van kieuwen, verdwijnen I, II en V aortakragen volledig, alleen de IIIrd, IVth en Vlth aortakrachten zijn aanwezig. De laterale dorsale aorta tussen de derde en vierde aortaboog (ductus caroticus) verdwijnt ook. Aldus, wordt de derde aortaboog samen met een deel van de ventrale aorta de halsslagader die zuurstofrijk bloed aan het hoofdgebied draagt.
de vierde aortaboog langs de laterale dorsale aorta vormt de systemische boog. De zesde aortaboog wordt de pulmocutane boog die veneus bloed levert aan longen en huid. De ductus arteriosus verdwijnt tijdens metamorfose. Volwassen anuranen hebben dus alleen III, IV en VI aortabomen. Deze worden ook behouden door vruchtwaterpunctie.
Reptilia:
bij reptielen worden de kieuwen volledig vervangen door longen. Alleen III, IV en VI aortabomen zijn aanwezig. Met de gedeeltelijke scheiding van het ventrikel in twee delen, het distale gedeelte van de conus arteriosus en de gehele ventrale aorta worden gesplitst in drie vaten, dat wil zeggen, twee aorta of systemische en een pulmonale. Rechter systemische boog (IV) ontstaan uit de linker ventrikel die zuurstofrijk bloed naar de halsslagader (III). De linker systemische (IV) en pulmonale aortae (VI) nemen hun oorsprong uit de rechter ventrikel. De linker systemische draagt gedeoxygeneerd of gemengd bloed naar het lichaam via dorsale aorta.
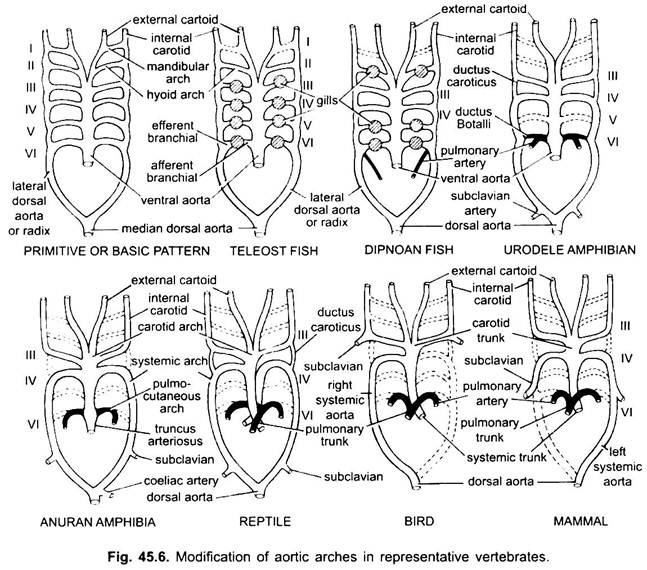
terwijl de longslagader gedeoxygeneerd bloed naar de longen voert. De ductus caroticus verdwijnt, maar blijft bestaan bij slangen en sommige hagedissen (Uromastix). De ductus arteriosus verdwijnt bij de meeste reptielen, maar blijft in verminderde vorm bestaan bij Sphenodon en sommige schildpadden. Door het mengen van bloed zijn reptielen koudbloedige dieren zoals vissen en amfibieën.
Aves:
bij vogels zijn de aortabomen III, IV en VI aanwezig. Ze volgen het algemene patroon van reptielen met enkele verschillen. Met de volledige verdeling van het ventrikel in twee delen, hebben de conus arteriosus en ventrale aorta gesplitst om twee schepen te vormen, systemische aorta die uit het linker ventrikel en een pulmonale aorta uit het rechter ventrikel.
derde aortaboog met restanten van de laterale en ventrale aortae vormt de halsslagader die uit de systemische aorta ontstaat. Vierde aortaboog vormt de systemische aorta alleen aan de rechterkant. Het verenigt zich met de laterale aorta van zijn eigen zijde en vormt de dorsale aorta. Een deel van de vierde aortaboog van de linkerkant vormt de linker subclavia slagader, de rest samen met de laterale dorsale aorta verdwijnt. De zesde aortaboog vormt de longaorta. Ductus caroticus en ductus arteriosus verdwijnen.
Mammalia:
bij zoogdieren blijven ook de III, IV en VI aorische bogen bestaan. Het ventrikel is volledig verdeeld in twee delen.
de conus arteriosus en ventrale aorta splitsen zich in twee bloedvaten:
(i) een systemische aorta die uit het linker ventrikel komt, en
(ii) een pulmonale aorta uit het rechter ventrikel. Derde aortaboog met resten van laterale en ventrale aortae vormt de halsslagader.
vierde aortaboog vormt alleen aan de linkerkant de systemische aorta, terwijl aan de rechterkant het proximale gedeelte een innominate en rechter subclaviale slagader vormt, de rest samen met de laterale dorsale aorta verdwijnt. De zesde aortaboog vormt de longaorta. De ductus arteriosus degenereert, maar blijft in sommige tot het uitkomen of geboorte in een verminderde vorm aan de linkerkant als een dunne ligamentum arteriosum.
Veneus Systeem:
Embryonale Aderen:
bij alle gewervelde embryo ‘ s is het veneuze systeem eenvoudig en vergelijkbaar. De nerven zijn meestal gekoppeld en symmetrisch gerangschikt. Bij embryo ‘ s zonder dooierzak wordt een sub – intestinale ader gevormd in het splanchnische mesoderm onder de darm.
het loopt rond de anus en wordt achteraf voortgezet als:
(i) een caudale ader in de staart. Bij alle embryo ‘ s met een dooierzak (al dan niet met dooier) ontstaat uit de dooierzak een paar vitelline-aderen die samenvloeien met het achterste deel van het zich ontwikkelende hart dat de sinus venosus wordt. In feite is de fusie van deze vitelline aderen verantwoordelijk voor de hartvorming bij benige vissen, reptielen en vogels. Elke vitellineader aan het achterste uiteinde sluit zich aan bij de subintestinale ader die op dezelfde manier is gevormd als bij de embryo ‘ s zonder dooierzak.
(ii) tussen de nieren ontstaat een paar subcardinale aderen en sluit zich aan bij de caudale ader.
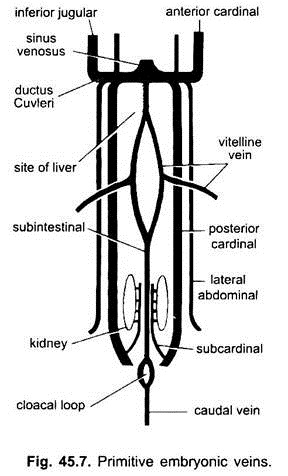
(iii) gepaarde voorste en achterste kardinale aderen worden gevormd, die het bloed uit het hoofd en achterste delen van het lichaam respectievelijk dragen. De voorste en achterste hoofdnerven van elke zijde verenigen zich tot een ductus Cuvieri of gemeenschappelijke hoofdnerf die naar binnen gaat door het transversale septum om de sinus venosus binnen te gaan.
bij vissen en salamanders (urodeles) komt een inferieure halsader uit de ventrale kant van de kop om zich aan te sluiten bij de gewone hoofdader. Het heeft geen homoloog in andere gewervelde dieren. In een vruchtwaterpunctie komt een paar laterale of ventrale abdominale aderen uit de lichaamsmuur om de gemeenschappelijke kardinale aderen binnen te gaan.
vissen:
gemeenschappelijke hoofdader (kanaal van Cuvier) komt van elke zijde in de sinus venosus en wordt gevormd door de fusie van anterieure en posterieure kardinalen. Bloed uit het hoofd wordt verzameld door de voorste kardinalen, en bloed uit de nieren en geslachtsklieren wordt verzameld door de achterste kardinalen.
in de kanalen van Cuvier komen ook de gepaarde laterale abdominale aderen die bloed uit de lichaamswand en gepaarde aanhangsels ontvangen. Het renale portaalsysteem bestaat uit de caudale ader en twee renale portale aderen die lateraal aan de nieren liggen en in de nieren capillariseren.
Leverportaalsysteem voert bloed uit de maag en darm en brengt het terug naar de lever, waar het, nadat het door een reeks sinusoïden is gegaan, via gepaarde hapatische aderen in de sinus venosus komt.
bij teleost ontbreken de laterale abdominale aderen, zodat het bloed van de subclavianen, dat de borstaanhangsels draineert, rechtstreeks in de sinus venosus terechtkomt, en het bloed van de iliacale aderen, dat de bekkenaanhangsels draineert, in de ansichtkaarten terechtkomt.
bij dipnoanen is één ventrale abdominale ader aanwezig, vermoedelijk afkomstig van een fusie van laterale abdominen. Deze ader ontvangt bloed van de ilacs door middel van gepaarde bekkenaders en komt in de rechter kanaal van Cuvier. Bovendien, vanuit het juiste postcardinale systeem, een nieuwe ader, maakt de postclaval zijn verschijning die van groot belang is bij hogere gewervelde dieren. Het is verbonden met de caudale ader en gaat door de lever naar de sinus.
bij embryo ‘ s van vruchtwaterpunctie zijn de laterale abdominale aderen bekend als navel-of allantoïsche aderen omdat ze de allantois afvoeren. De vitelline en naveladers gaan bij de geboorte verloren, alleen de restanten ervan blijven bestaan.
wijzigingen van aderen:
zoals bij slagaders zijn de aderen van verschillende gewervelde dieren gerangschikt volgens hetzelfde basisplan. Elke door hen getoonde variatie volgt een sequentie in de gewervelde reeks. In zijn ontwikkeling gaat het veneuze systeem van hogere gewervelde dieren door de stadia die in embryo ‘ s van lagere vormen worden gezien.
1. Wanneer de lever wordt gevormd het proximale deel van de vitelline aderen of subintestinale aderen vormt de lever aderen tussen de lever en het hart, het distale deel van de linker vitelline ader of subintestinale ader wordt de lever portale ader die sinusoïden in de lever vormen om te leiden tot een lever portaal systeem aanwezig in alle.
2. Anterior kardinaal aders blijven bestaan als interne halsader.
3. Behalve in vissen, worden de gemeenschappelijke hoofdnerven de precavale aderen die de sinus venosus (amfibieën, reptielen) of de rechteroorschelp (vogels, zoogdieren) binnendringen. In elke voorpoot wordt een subclavische ader gevormd die zich aansluit bij de precavale ader.
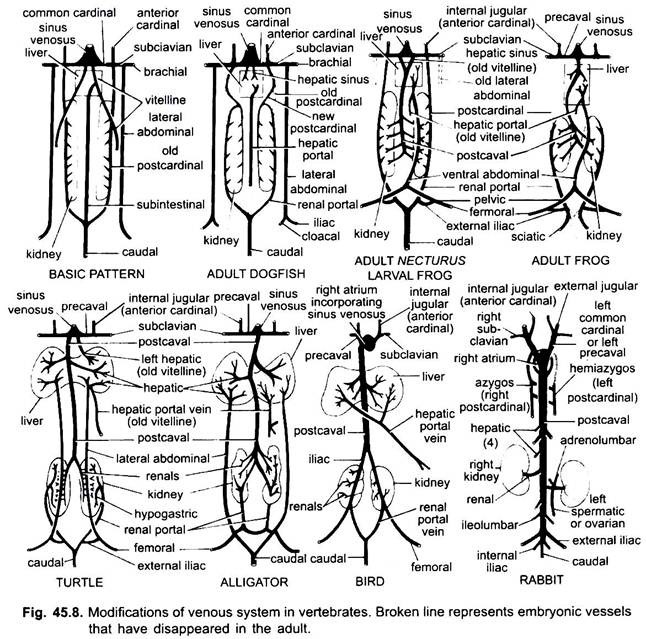
4. De caudale ader verliest zijn verbindingen met de subintestinale en subcardinale aderen, het voorste deel splitst zich in twee takken die de achterste kardinale aderen bij alle behalve zoogdieren verbinden.
5. Bij vissen blijven de achterste kardinale aderen als zodanig bestaan, maar in andere delen splitsen ze zich in twee delen, een voorste gedeelte dat bij amfibieën, reptielen en vogels verdwijnt, maar bij zoogdieren vormt het rechter voorste gedeelte een azygosader, de linker kan een hemiazygosader vormen die zijn verbinding met de precavale verliest en de azygosader door een transversale anastomose verbindt; de achterste delen van de achterste kardinale aderen die zich bij de caudale ader hebben aangesloten, worden de renale portale aderen.
de nierportaalader van elke zijde vormt haarvaten in de nier tot een nierportaalsysteem dat volledig is bij vissen, amfibieën en reptielen, maar bij vogels sterk afneemt en afwezig is bij zoogdieren omdat de achterste delen van de achterste hoofdader verdwijnen bij zoogdieren.
6. De renal portal aders van vissen verzamelen bloed alleen van de staart, maar in amfibieën, reptielen en vogels ze ook usurpped de aders van de benen.
7. Bij sommige vissen en alle tetrapoda sluiten de vitelline-aders zich aan in de subcardinale ader en vormen zo een postcavale ader. Bij amfibieën en reptielen strekt de postcavale ader zich uit tot de achterste uiteinden van de nieren, bij vogels sluit hij zich aan bij de renale portale aderen, waardoor het renale portale systeem wordt verminderd, bij zoogdieren sluit de postcavale ader zich aan bij de aderen van de benen en de staart, zodat het renale portale systeem volledig wordt geëlimineerd.
8. De twee laterale abdominale aderen blijven als zodanig in vissen, maar van dipnoi omhoog versmelten ze tot een voorste abdominale ader die de hepatische poortader in de buurt van de lever verbindt, de voorste abdominale ader, dus, verbindt de nier portaal en lever portaal systemen. Een iliacale ader wordt gevormd in elke achterpoot die de voorste buikader verbindt. De voorste buikader begint zijn belang bij reptielen te verliezen, bij vogels wordt het gewijzigd om de epigastrische en coccygeo-mesenterische aderen te vormen, bij zoogdieren verdwijnt het, behalve bij Tachyglossus.
9. In lucht-ademende vormen, long aders groeien uit de linker oorschelp en in de longen.
10. De navelstreng (allantoic) aderen van het embryo verdwijnen wanneer de longen functioneel worden als ademhalingsorganen.
Tetrapoda:
het veneuze systeem van amfibieën lijkt sterk op dat van longvissen, behalve dat de abdominale ader het hepatische portaalsysteem binnendringt in plaats van de sinus venosus. Anterieure kardinalen blijven bestaan als interne halsader in alle volwassen tetrapoden. Inferieure halsslagaders zijn afwezig. De gewone kardinalen worden de vena cavae of precavals, die samengaan met sinus venosus bij amfibieën en reptielen.
bij vogels en zoogdieren komen precavals rechtstreeks in de rechter oorschelp van het hart, omdat sinus venosus afwezig is. In de mens, kat, enz., linker precaval is verloren, zodat het bloed van de linkerkant de rechter precaval via een brachio-cephalic tak binnenkomt. Bij longloze salamanders zijn de longaders afwezig omdat het interatriale septum onvolledig is.
de postcardinale aderen sluiten zich aan bij de caudale ader posterior en precavals anteriorly om de gemeenschappelijke kardinalen te vormen bij larvekikker en Necturus.
bij reptielen is er een grotere ontwikkeling van pulmonale aderen en postcavale aderen en een afname van het belang van het nierportaalsysteem dat bloed naar de nieren brengt vanuit het achterste deel van het lichaam.
bij vogels zijn er twee functionele precavale aderen gevormd door de Vereniging van de halsader en het subclavium aan elke kant, en een volledige post-cavalerie. Het ontvangt bloed van de ledematen door middel van de renale portalen, die door de nieren, maar niet breken in haarvaten en zijn niet vergelijkbaar met de renale portalen van lagere gewervelde dieren.
bij zoogdieren kunnen er één of twee precavale aderen zijn. Er is een enkele postcavale ader, waarvan de embryologische ontwikkeling zeer gecompliceerd is. Alle aderen van de cavalerie komen direct in de rechter oorschelp terecht, omdat de sinus venosus in het embryonale leven in de wand van deze hartkamer wordt opgenomen. Er is geen nierportaalsysteem, hoewel het leverportaalsysteem sterk lijkt op dat van andere gewervelde dieren.