PUBLICIDADE:
neste artigo vamos discutir sobre o sistema digestivo de vertebrados, com a ajuda de diagramas.
tracto digestivo embrionário:
Archenteron:
o archentron embrionário torna-se o revestimento do tracto digestivo adulto e de todos os seus derivados. O mesoderme splancnico adiciona camadas de tecido conjuntivo e músculos lisos ao redor do arcentro. A invaginação ectodérmica da cabeça forma o estomodaeum levando à cavidade oral, e uma invaginação ectodérmica semelhante forma proctodaeum, que leva ao fundo. Publicidade:
o estomodeu torna-se a cavidade bucal adulta e dá origem a esmalte dentário, cobertura epitelial da língua, glândulas, por exemplo, mucosas, veneno e salivar, etc., e a bolsa de Rathke da glândula pituitária anterior. O proctodaeum forma uma pequena parte terminal da cloaca em vertebrados inferiores e recto em mamíferos.
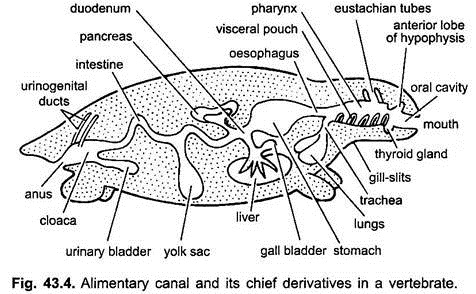
O canal alimentar em embriões a partir do estômago para cloaca é anexado a dorsal do corpo parede por uma dobra do peritônio, chamado o mesentério dorsal e ventral do corpo parede por um mesentério ventral. Em adultos, o mesentério dorsal persiste, mas o mesentério ventral desaparece deixando apenas na região do fígado e bexiga urinária.
Aparelho Digestivo de adulto: O trato digestivo diferencia para diferentes funções nas seguintes regiões: boca, cavidade bucal, faringe, esófago, estômago, intestino delgado, intestino grosso e cloaca. Na sequência de crescimentos surgem a partir do trato digestivo – glândulas orais, bolsa de Rathke, glândula tiroideia, fissuras nas guelras, cavidade timpânica, timo e outras glândulas de fissuras nas guelras, traqueia, pulmões, bexiga de banho, fígado, pâncreas, saco de gema e bexiga urinária. Publicidade:
Histologia:
a parede do canal alimentar é feita de quatro camadas concêntricas.
São:
(i) um peritoneu visceral externo ou revestimento seroso é feito de células mesoteliais e fina camada de tecido conjuntivo. Falta no esófago,
publicidade:
(ii) Abaixo está uma camada muscular formada por fibras musculares lisas dispostas em fibras musculares longitudinais exteriores e circulares interiores. Entre as duas camadas de músculos está uma rede de células nervosas e fibras nervosas do sistema nervoso autônomo, conhecido como plexo myentérico ou plexo de Auerbach.
(iii) Abaixo da camada muscular é um submucosa feito de tecido conjuntivo ter fibras elásticas, gordura, vasos sanguíneos e linfáticos, células nervosas e fibras glândulas,
(iv) A camada mais interna é uma mucosa composta por três regiões:
(a) Externo-mais estreito musculares mucosa do exterior longitudinal interna e circulares suaves fibras musculares.
(b) Meio fina camada de lâmina própria de tecido conjuntivo, vasos sanguíneos, nervos e nódulos de tecido linfático e
(c) Uma membrana basal apoio de uma camada de células epiteliais colunares, que são, muitas vezes, glandular e ciliadas. A boca é a abertura que conduz à cavidade bucal. Em lampreias (ciclostomas) é uma abertura circular na base do funil bucal e permanece permanentemente aberta devido à falta de Mandíbulas, etc. Em gnatostomos é terminal. A boca é limitada por lábios que são imóveis e formados de pele cornifiada em peixes, anfíbios e répteis. Em mamíferos estes são carnudos e musculados. Relação das vias nasais com cavidades bucais em diferentes vertebrados cavidade bucal:
publicidade:
o espaço entre os lábios e as mandíbulas é um vestíbulo. Pode ser limitado por fora pelas bochechas e por dentro pelas gengivas. As glândulas mucosas das bochechas abrem-se no vestíbulo. A boca se abre em uma cavidade bucal, que é um espaço entre a boca e a faringe. O ponto exato onde o ectoderme estomodeal e endodermia faríngea se fundem é variável e não é fácil de discernir.
em elasmobrânquios e na maioria dos peixes ósseos, as cavidades nasais não se abrem na cavidade bucal. Em Chondrichthyes e tetrapoda (anfíbios e a maioria dos répteis) o das cavidades nasais abrem na cavidade bucal por choanae ou interno narinas, que são primitivamente colocado anteriormente, mas em crocodilos, aves e mamíferos que se tornar posterior da faringe devido à formação de um palato secundário, o que efetivamente separa o respiratória nasal passagem da boca da cavidade, ou a passagem do alimento.
em aves, este palato é fissura devido à qual cavidades nasais e bucais se comunicam entre si. Nos mamíferos, o palato secundário é mantido posteriormente como um palato mole membranoso. Em seres humanos, o palato macio está preso na faringe na forma de processo carnudo, chamado uvula.
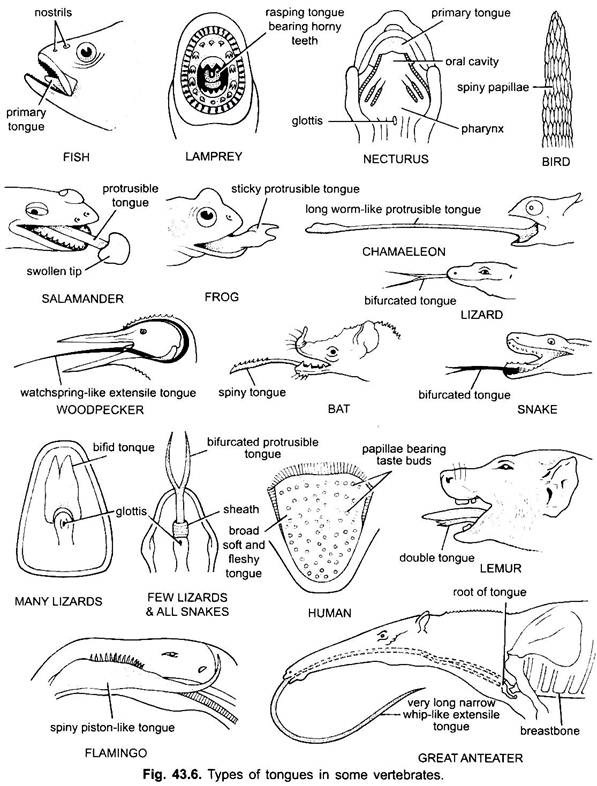
Derivados da Cavidade Bucal:
1. Glândulas orais:
existem dois tipos de glândulas multicelulares integrais que se abrem na cavidade bucal. São glândulas mucosas e glândulas enzimáticas. Peixes e anfíbios aquáticos têm apenas glândulas mucosas. Os répteis têm glândulas em grupos, tais como Palatino, lingual, sublingual e glândulas labiais chamadas de acordo com a localização, eles também produzem muco. Em cobras venenosas, as glândulas labiais superiores são modificadas para segregar veneno, enquanto no monstro de Gila as glândulas sublinguais produzem veneno. As aves têm glândulas sublinguais e uma glândula no ângulo da boca. Os mamíferos têm muitas glândulas mucosas pequenas, além de glândulas salivares verdadeiras e ampliadas, que são enzimáticas. São glândulas salivares parótidas, sublinguais, submaxilares e infraorbitais, secretando mucina e ptialina.
2. Língua:
a língua é encontrada principalmente em todos os vertebrados. A língua em vertebrados mostra muita diversidade e não são homólogos. Nos ciclostomos, há uma língua musculosa, carnuda, rasgando com dentes córneos para rasgar a pele e músculos de sua presa.
os peixes têm uma língua primária formada por uma dobra carnuda do chão bucal. Não tem músculos, mas receptores e dentes estão presentes na língua em alguns peixes ósseos. A língua está coberta com membrana mucosa. Em alguns anfíbios a língua é inexistente ou inamovível. A maioria dos anfíbios, no entanto, tem uma língua protrusível e em algumas rãs e sapos pode ser dobrado de volta sobre si mesmo quando não está em uso.
pode ser expelida para fora da boca por um rápido fluxo de linfa para a captura dos insectos. A língua em lagartos e cobras é muitas vezes altamente desenvolvida. Em camaleões, é muito extensível usado para capturar insetos. A ponta é espessa e pegajosa. A ponta bifurcada da língua nas cobras serve como um meio de transferir estímulos químicos do ambiente externo para os órgãos vomero-nasais emparelhados no céu da boca. Em tartarugas e crocodilianos, a língua não pode ser estendida. A língua amniota tem músculos voluntários, recebe o nervo hipoglosso e tem glândulas e papilas gustativas. Também desenvolve músculos intrínsecos que movem a língua. Em pássaros, a língua é esbelta e tem uma cobertura excitada. Em algumas aves a língua é imóvel, enquanto em algumas aves é longa, protractil e muitas vezes usado para capturar a comida.
na maioria dos mamíferos, exceto baleias, a língua é altamente desenvolvida e capaz de movimentos consideráveis, além de extensão e retração, devido à presença de uma série de músculos intrínsecos.
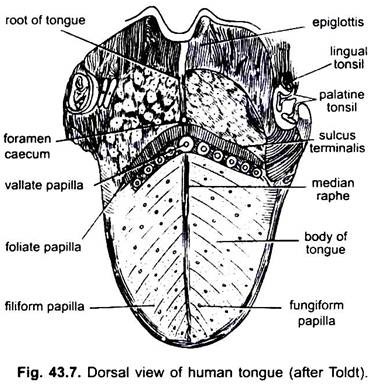
em mamíferos, a membrana mucosa abaixo da língua forma uma dobra mediana, chamada frenulum que une a língua ao chão da boca. Em mamíferos, a superfície superior da Língua apresenta quatro tipos de papilas, (filiforme, fungiforme, foliar e circunvalado), com papilas gustativas exceto as papilas filiformes.
3. Dentes:
vertebrados têm dois tipos de dentes ligados aos ossos da mandíbula – dentes epidérmicos e dentes verdadeiros. Os dentes epidérmicos são melhor desenvolvidos em ciclostomas. São estruturas duras, cónicas e excitadas derivadas do estrato córneo. Em lampreias eles são encontrados nas paredes do funil bucal e na língua. Larvas de sapos e sapos têm dentes epidérmicos serrilhados em filas nos lábios. Em mamíferos, o patifo adulto tem dentes epidérmicos. Dentes verdadeiros: dentes verdadeiros: dentes verdadeiros: dentes verdadeiros: dentes verdadeiros: dentes verdadeiros não são encontrados em baleias-de-barbatanas e comatores em mamíferos, e agnatões, esturjões, alguns sapos, sirenes, tartarugas e aves modernas, etc. Em vertebrados inferiores (como peixes, anfíbios e a maioria dos répteis) os dentes podem ser substituídos continuamente por um número indefinido de vezes, tais dentes são chamados de polifiodonte. Estes dentes são homodont (tipo similar) e acrodont. (com os ossos do maxilar). Na maioria dos mamíferos, os dentes são difiodonte, tecodonte e heterodonte. Em alguns mamíferos estes são monofiodontes com apenas um conjunto de dentes, por exemplo, moles, Esquilo Indiano. Os dentes são semelhantes em estrutura às escamas placóides dos tubarões, formados por uma cavidade central de polpa, em torno da qual está presente uma camada espessa mas macia, a dentina, que é coberta externamente por um esmalte fino e extremamente duro. Estes são supostos ter derivado de escamas ósseas de ostracodermes e placodermes. Para mais detalhes, os leitores podem ver a dentição em mamíferos.
4. Adenohypophysis:
o lobo anterior da glândula pituitária desenvolve-se como uma evaginação dorsal do estomodeu, chamada bolsa de Rathke, que se contrai para formar os lobos anterior e médio da glândula pituitária (adenohypophysis). O lobo posterior da hipófise ou neurohypophysis é a evaginação ventral de diencéfalo, chamado de infundibulum. Assim, é a parte nervosa.
faringe:
a parte do canal alimentar imediatamente atrás da cavidade bucal é uma faringe, forrada com endodermia. É uma passagem comum servindo tanto para a digestão e respiração. Como parte do sistema digestivo é usado como uma passagem para alimentos da cavidade bucal para o esôfago, seus músculos iniciam a deglutição.
em peixes, a faringe é grande e lateralmente perfurada para fendas de gill, enquanto em tetrapoda, é curta e tem aberturas de narinas. Em embriões, a parede da faringe emite uma série de evaginações que se desenvolvem em espiráculos, fissuras em guelras, bexigas de ar, pulmões, amígdalas e algumas glândulas endócrinas (por exemplo, timo, tiróide e paratiróides).
esófago:
o esófago é curto na maioria dos peixes e anfíbios por falta de pescoço, mas é mais longo em amniotas devido à presença de pescoço. O esófago dos répteis é mais alongado do que o dos peixes e anfíbios. Em aves granívoras e carnívoras, uma porção do esófago é ampliada em uma bolsa semelhante a saco chamada Safra, que serve para armazenar alimentos que foram consumidos rapidamente.
a cultura é essencialmente carente de glândulas digestivas, embora nos pombos a cultura tenha 2 glândulas vegetais em ambos os sexos, eles não são realmente glândulas, mas estruturas de formação de células, as células formam “leite de pombo” que é alimentado para os jovens. Em mamíferos, o esófago é longo, carece de glândulas e varia em relação ao comprimento do pescoço.
ele passa pelo diafragma, a porção abaixo do diafragma é coberta com peritoneu visceral que está faltando a partir da parte superior. O esófago tem glândulas mucosas. Seu forro forma dobras longitudinais, ou papilas carnudas (elasmobranchos) ou papilas excitadas em tartarugas marinhas. Histologicamente, o esófago difere do resto do canal alimentar em três factos:
(i) não tem peritoneu visceral porque se encontra fora do coeloma, a sua camada exterior de cobertura é uma fina túnica adventícia.
(ii) As fibras musculares na sua parte anterior são listradas, a parte média tem ambos músculos listrados e não listrados, e a parte posterior tem apenas músculos não listrados. Mas há exceções em mamíferos que ruminam, todos os músculos são listrados ou voluntários.
(iii) O revestimento da membrana mucosa é feito de células epiteliais escamosas estratificadas e não de células colunares.
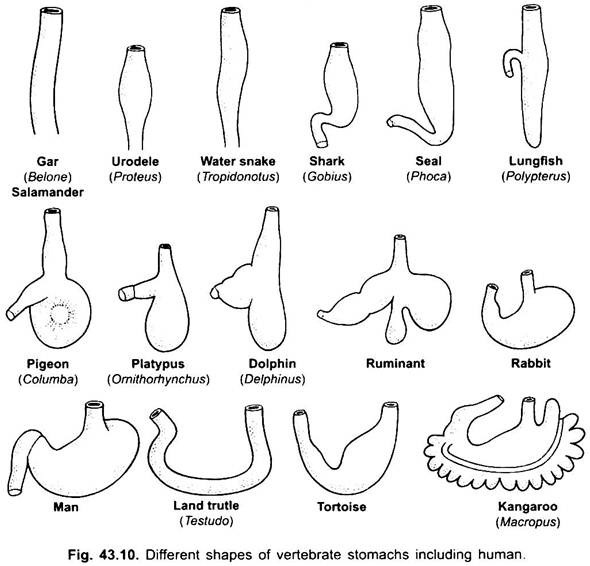
Estômago:
não Há praticamente nenhum estômago em cyclostomes, de quimeras, de pulmão, peixes e alguns primitivo peixes teleósteos de cerca, uma vez que não tem glândulas gástricas, mas a maioria dos peixes e tetrapoda é dilatado para o armazenamento e a maceração de alimentos sólidos, e a digestão do alimento, pois contém glândulas gástricas.
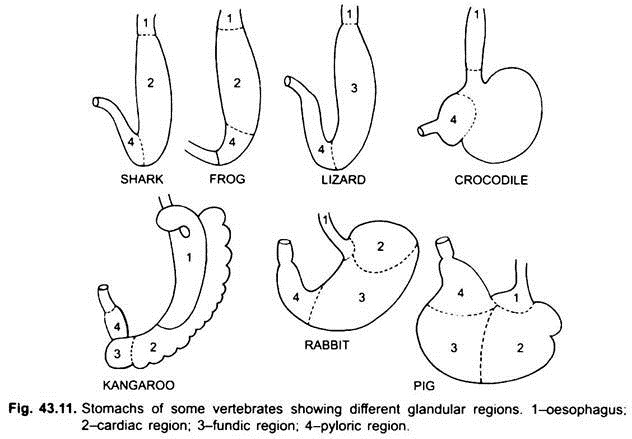
A primeira parte do estômago, junto ao esófago, é a cardíaca região e a extremidade inferior perto do intestino é a região pilórica, que tem um piloro ou válvula pilórica em que a mucosa de revestimento é cercado por uma espessa músculo do esfíncter que regula a abertura e o fechamento da pilórica do estômago para o intestino.
o estômago é recto nos ciclostomas, gar, Belone, etc., e em forma de fuso em Proteus, Necturus, alguns lagartos e cobras. Em tartarugas e tartarugas, é um grande tubo curvado, e em elasmobranches o estômago é em forma de J. Em crocodilos e aves o estômago tem duas partes, um proventriculo com glândulas gástricas, e uma moela altamente muscular, que representa a região pilórica e tem um revestimento duro e cornificado para moer alimentos.
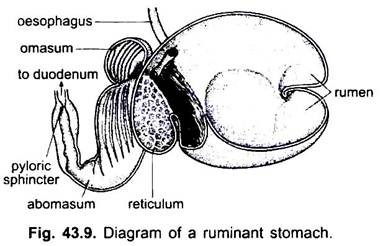
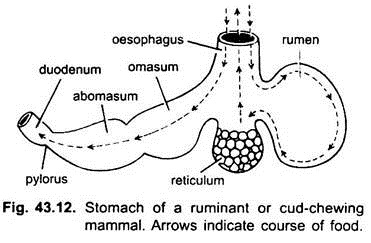
nos mamíferos o estômago encontra-se transversalmente e pode ser um saco simples ou dividido em 3 regiões, nomeadamente cardíaca, fundica e pilórica e cada região tem as suas glândulas gástricas. Em muitos ruminantes o estômago tem quatro câmaras-um rúmen, reticulum, omasum e um abomasum. Alega-se que as três primeiras câmaras são modificações do esôfago, e abomaso é o verdadeiro estômago representando as partes cardíacas, fundas e pilóricas do estômago. Foi demonstrado embriologicamente que todas as quatro câmaras são regiões modificadas do estômago. Em camelos, não há omasum, o rúmen e reticulum têm células de água como bolsa que foram acreditados armazenar água, mas eles são provavelmente digestivos. Histologicamente, o estômago tem as partes típicas do canal alimentar, mas tem duas peculiaridades, a mucosa muscular é feita de uma camada longitudinal exterior e uma camada circular interna de músculos. O revestimento do epitélio é espesso com vários tipos de células glandulares formando glândulas gástricas de três tipos chamados glândulas gástricas cardíacas, fundicas e pilóricas.
as glândulas cardíacas e pilóricas segregam apenas muco das suas células superficiais. As glândulas fundicas (ou glândulas cardíacas em alguns) têm três tipos de células, as células mucosas do pescoço produzem muco, as células oxinticas produzem ácido clorídrico, eles podem estar presentes na região cardíaca também, as células zymogen ou células pepticas secretam pepsina.
na maioria dos animais, as células de zimogénio segregam também duas proenzimas chamadas propepsina e pró-Nina, que são convertidas pelo ácido clorídrico em pepsina e rennina, respectivamente. As secreções de todas as células do estômago formam uma mistura chamada sumo gástrico.
Intestino Delgado: O intestino delgado é longo, estreito e enrolado após o piloro. É a parte mais importante do trato digestivo porque a digestão e absorção de alimentos ocorrem nele. Nos ciclostomas, o intestino é um tubo curto e recto com uma aba longitudinal espiralmente arranjada que se estende até ele.
em elasmobranchos, é dividido em pequenas e grandes porções, e a pequena porção tem uma válvula espiral que aumenta grandemente a superfície absorvente. Uma válvula espiral também está presente no intestino delgado de alguns peixes ósseos mais primitivos, mas está faltando em formas mais altas em que o intestino é longo e enrolado.
In caecilians, it is little coiled and not differented into a small and large tract. Em sapos e sapos é relativamente longo e enrolado. Em répteis, é mais enrolado do que em anfíbios. Pela primeira vez em vertebrados um ceco ou divertículo cego surge na junção de pequenos e grandes intestinos. No entanto, isto não é permanente em todos os répteis. Em aves, o intestino delgado é enrolado ou enrolado e uma ou duas cólicas também estão presentes na junção de intestinos pequenos e grandes. Na maioria dos mamíferos também o intestino delgado é proporcionalmente longo e enrolado. Sua duração, no entanto, está correlacionada com os hábitos alimentares. Em herbívoros é relativamente mais longo em comparação com insectívoros e carnívoros.
Há um cego bolso ou ceco-se na junção do cólon e do intestino delgado, que é geralmente pequena em espécies carnívoras e bastante longo, em muitos herbívoros. A primeira parte do intestino delgado é o duodeno, que é o início curto a partir de piloro e termina além da entrada de ductos pancreáticos e hepáticos.
tem muitas vilosidades dobradas e contém glândulas de Brunner ramificadas na submucosa que segregam muco, alguns fluidos aquosos alcalinos e uma pequena enzima. O duodeno também produz duas hormonas denominadas secretina e colecistocinina que estimulam o pâncreas e a vesícula biliar a libertar os seus sumos. Condutas da vesícula biliar e pâncreas abrem-se no duodeno.
atrás do duodeno está um íleo,que apenas em mamíferos é diferenciado em um jejuno menor anterior e íleo mais longo posterior. Um grande número de glândulas digestivas pequenas estão presentes no intestino delgado. Elas são glândulas tubulares ou criptas de Lieberkuhn encontradas através de todo o comprimento, elas secretam muco e uma succus entericus que tem várias enzimas.
o revestimento do intestino delgado é dobrado para formar vilosidades pequenas, o que aumenta a área superficial para secreção e absorção. As vilosidades são cobertas densamente por minúsculas projeções tipo dedo, chamadas microvilli que ajudam na absorção para as vilosidades. Em mamíferos nódulos de tecido linfóide chamados manchas de Peyer são encontrados no íleo.
Intestino grosso:
intestino grosso tem um diâmetro maior do que o intestino delgado. É geralmente curto em peixes, anfíbios, répteis e aves, mas em mamíferos é longo. Em formas mais baixas, o intestino grosso forma um recto, mas em tetrapoda tem um cólon e um recto terminal. Na maioria dos peixes e anfíbios, a parte terminal do rectum leva a uma cloaca formada pelo proctodaeum.
o recto, as condutas de excreção e os canais genitais abrem-se na cloaca e abrem-se para o exterior por uma abertura cloacal. Mas em muitos peixes ósseos e todos os mamíferos (exceto monotremados), as condutas recto e urinogenital têm aberturas separadas para o exterior.; a abertura do primeiro é um ânus.
o recto de mamíferos não é homólogo com o recto de vertebrados, uma vez que nos mamíferos é derivado pela separação da cloaca embrionária. Na maioria dos embriões de vertebrados há um intestino pós-anal como uma extensão do intestino na cauda, mas ele desaparece mais tarde.
em elasmobrânquios, o intestino grosso apresenta um par de glândulas rectais que secretam muco e cloreto de sódio. Em amniotes há uma válvula ileocólica na junção de pequenos e grandes intestinos, que está ausente em peixes. Impede que as bactérias entrem no íleo a partir do cólon.
in amniotes from this junction arises an ileocolic caecum which is two in birds. Contém bactérias digestoras de celulose. É muito longa em mamíferos herbívoros (coelho, cavalo, vaca, etc.). Em primatas, o ceco é pequeno, com um apêndice vermiforme vestigial.
glândulas digestivas:
1. Fígado:
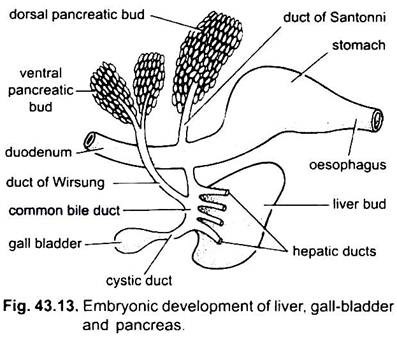
o fígado surge como um único ou duplo crescimento a partir da parede ventral do arcentro embrionário. Este crescimento forma um divertículo hepático oco, que logo se diferencia em uma parte anterior, que prolifera para se tornar o fígado e seus ductos biliares, e uma parte posterior, o que dá origem à vesícula biliar e ducto cístico. Os dutos biliares unem-se para formar um duto hepático que se une com o duto cístico para formar um duto biliar comum ou duto choledocus. A região do arquentério a partir do qual o fígado Surge torna-se o duodeno. O fígado é a maior glândula lobada do corpo, suspensa por uma camada dupla de peritoneu do septo transversal ou seu representante.
Uma vesícula biliar é para o armazenamento da bile, secretada pelas células hepáticas, encontra-se no fígado e drena para o duodeno através do ducto biliar comum formado pela união do ducto cístico e do ducto hepático. Uma vesícula biliar não é indispensável e está faltando em muitas aves e mamíferos.
o fígado está presente em todos os vertebrados. Em ciclostomos, é pequeno, com um só lóbulos (lampreias) e dois lóbulos em hagfishes. É bilobada em elasmobranches, duas ou três lóbulos em peixes ósseos, anfíbios, répteis e aves e muitos lobados em mamíferos. O fígado é longo, estreito e cilíndrico em peixes, urodeles e cobras. É curto, largo e achatado em aves e mamíferos. Uma vesícula biliar e ducto biliar estão presentes nos ciclostomas larvares, mas eles estão ausentes no adulto. Peixes, anfíbios e répteis geralmente têm uma vesícula biliar, mas falta em muitas aves. A maioria dos mamíferos tem uma vesícula biliar, mas está ausente em Cetácea e Ungulata.
o fígado secreta uma bílis aquosa e alcalina, mas não tem enzimas. Neutraliza a acidez dos alimentos que entram no duodeno. Ajuda na digestão de gorduras.
2. Pâncreas:
o pâncreas é formado a partir do endodérmico do arquentrão embrionário. Um único divertículo dorsal do duodeno embrionário e um ou dois rebentos ventrais do fígado formam divertículo pancreático. As partes proximais da diverticula formam condutas pancreáticas, mas estas sofrem redução ou fusão de modo que apenas uma ou duas condutas pancreáticas permanecem no adulto, elas abrem-se no duodeno separadamente ou depois de se unirem com o ducto biliar comum.
as partes distais da divertículo sofrem a formação da massa principal das células pancreáticas às quais se adicionam derivados mesodérmicos. Assim, uma única glândula é feita que tem vários lóbulos formando um pâncreas difuso ou compacto. O pâncreas é uma glândula exócrina e endócrina, ligada por fios delicados de tecido conjuntivo. A parte exócrina segrega enzimas digestivas que são derramadas no duodeno através de condutas pancreáticas. Enquanto o endócrino segrega hormonas como a insulina e o glucagon. O pâncreas está presente em todos os vertebrados. Em lampreias, alguns peixes ósseos, lungfishes e tetrápodes inferiores, é um órgão difuso embutido no fígado, mesentérias e parede intestinal. Os Hagfishes têm um pâncreas pequeno. Os elasmobranchos têm um pâncreas bem definido. Em tetrapoda superior é geralmente uma glândula compacta. Uma ou duas condutas pancreáticas abrem-se no duodeno.