- Introdução
- Resultados
- A DAP-Cre Driver de Marcas e É Limitado a TH-Expressando Noradrenérgico Regiões do Tronco cerebral Que São Inibidas pelo CNO de Administração
- CNO-hM4D mediou a perturbação dos neurónios noradrenérgicos em ratinhos adultos
- CNO-hM4D Perturbation of DBH-Cre Neurons Results in a Hypercapnic Deficit
- controlos habitados de irmãos injectados com 10 mg / kg de CNO revelaram um défice ventilatório Hipercapnico
- Habituados e Unhabituated tipo selvagem Animais Injetados Com Salina Não Mostrou Alterações Presaline e Postsaline
- as razões de concentração da clozapina para CNO são mais elevadas no cérebro do que no soro
- discussão
- materiais e métodos
- aprovação ética
- reprodução, fundo genético, e manutenção de ratinhos
- Histologia
- electrofisiologia
- Plethysmography
- Cromatografia Líquida-espectrometria de massa
- Recolha e Análise de Dados
- Pletismografia
- Estatísticas
- Pletismografia
- Cromatografia Líquida-espectrometria de massa
- Autor Contributions
- Funding
- Declaração de conflito de interesses
- agradecimentos
Introdução
redes neuronais que subjazem respiratório chemosensory reflexos são um alvo primário para a compreensão da etiologia de várias comportamentais e distúrbios fisiológicos. Perturbado chemosensory reflexos são a hipótese de jogar um papel no congênitas e adultos doenças, incluindo Síndrome de Morte Súbita Infantil (SIDS), Congênitas Central, Síndrome de Hipoventilação (CCHS), central de apnéias do sono, respiração anormal na Síndrome de Rett, obesidade e síndrome de hipoventilação (Obonai et al., 1998; Ozawa et al., 2003; Amiel et al., 2009; Ramanantsoa et al., 2011; Lavezzi et al., 2013). Sob a hipótese do falso alarme por asfixia, pensa-se que a ativação quimiossensorial inadequada ou a hipersensibilidade desempenham um papel em subconjuntos de pacientes que sofrem de transtorno de pânico (Klein, 1993). A disfunção quimiossensorial também pode desempenhar um papel em doenças neurodegenerativas através da respiração desordenada do sono, que está associada com progressão acelerada (Hakim et al., 2012; Verbraecken e McNicholas, 2013; Bahia e Pereira, 2015; Snyder et al., 2017). Assim, uma melhor compreensão das redes quimiossensoriais do tronco cerebral irá fornecer pistas importantes para uma série de patologias comportamentais e fisiológicas.
receptores de designer Farmacogenéticos activados exclusivamente por medicamentos de marca (DREADDs) (Armbruster et al., 2007) têm sido empregados em uma série de estudos para mapear as populações neurais no chemoreflex respiratório (resposta respiratória para níveis elevados de CO2 no sangue). DREADD tecnologia em combinação com intersectional genética de implantação, tem sido utilizado para silenciar altamente segmentados neuronal populações observando a função respiratória em consciente e desenfreada ratos pelo nosso laboratório e outros, evitando muitos confunde anteriores circuito de mapeamento de abordagens (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Estes e a maioria dos outros estudos quase sempre incluíram CNO apenas controles irmãos não-DREADD expressando que não mostraram efeitos quimiosensoriais ou outros efeitos respiratórios, argumentando que CNO não tinha efeitos fora do alvo na respiração em ratos conscientes e sem restrições, apesar da dose elevada utilizada, 10 mg/kg. No entanto, foi encontrado em vários outros estudos que o CNO e seus produtos de metabolismo de costas poderiam ter efeitos fora do alvo sobre o comportamento e locomoção em uma variedade de ensaios, mas a saída respiratória não foi abordada (Guettier et al., 2009; Joober, 2010; MacLaren et al., 2016; Gomez et al., 2017; Ilg et al., 2018; Mahler and Aston-Jones, 2018; Manvich et al., 2018; Padovan-Hernandez and Knackstedt, 2018). Foi também demonstrado que o CNO e os seus metabolitos não se distribuem de forma equivalente entre o sistema circulatório e o cérebro (Gomez et al., 2017).
a second, unaddressed concern arises from the stress induced by the experimental paradigm, including holding animals in a whole-body plethysmography chamber, handling, intra-peritoneal injection, and rectal temperature measure. Em estudos respiratórios anteriores do DREADD, ratos naïve foram introduzidos em uma câmara pequena (140-400 mls) e autorizados a aclimatar 20-40 min antes da aquisição de dados (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). No entanto, não é claro se esta quantidade de tempo é suficiente para minimizar as alterações respiratórias induzidas pelo stress que podem actuar como um factor de interacção com os efeitos da clozapina.
em nossos estudos para examinar o papel do sistema noradrenérgico (NA) no controle respiratório, procuramos empregar o RC bem estabelecido::P_hM4D DREADD allele (Ray et al., 2011) to test the role of NA neurons in baseline and hypercapnic respiration in unrestrained adult animals using whole-body plethysmography. Porque os neurónios noradrenalina e NA também desempenham um papel central nas respostas ao stress (Valentino e Van Bockstaele, 2008; Chen et al., 2018), realizamos uma série de estudos para comparar a habituação extensiva (múltiplas exposições ao paradigma experimental em que o animal aprende que a experiência não é ameaçadora ou sobrevivível) e alta vs. doses CNO baixas (1 mg / kg) com protocolos DREADD publicados anteriormente (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Aqui, mostramos pela primeira vez que doses sistémicas elevadas de CNO são capazes de provocar efeitos fora do alvo numa função respiratória autónoma em ratos conscientes. Nós também revelam que o fora-de-alvo efeito de CNO em chemosensory respiratória saída é efetivamente desmascarado por extenso habituação e não seria, portanto, aparente anteriores CNO estudos de controle que não acostumar os animais antes respiratória de medição, utilizando apenas um curto período de aclimatação antes da coleta de dados. Juntos, estes resultados sugerem que populações neuronais previamente mapeadas podem indiretamente afetar o controle respiratório através de papéis potenciais na regulação das respostas ao estresse. Nomeadamente, estes dados estão em consonância com relatórios recentes que sugerem que o CNO não é biologicamente inerte em doses elevadas através da conversão metabólica em clozapina (MacLaren et al., 2016; Gomez et al., 2017) e que os efeitos comportamentais fora do alvo podem se manifestar não só a partir da perturbação dos circuitos comportamentais, mas também de interrupções para circuitos autônomos subjacentes e homeostase.
Resultados
A DAP-Cre Driver de Marcas e É Limitado a TH-Expressando Noradrenérgico Regiões do Tronco cerebral Que São Inibidas pelo CNO de Administração
Para examinar a expressão e a especificidade do DAP-Cre linha, utilizou-se um único recombinase programa de criação (Figura 1A), em que cruzou o driver para o Ai9 linha (Madisen et al., 2010), que expressa um tdtomato floxado. A coloração com um anticorpo da tirosina hidroxilase (TH) revelou que a expressão do tdTomato no tronco cerebral estava limitada às regiões que expressavam O TH, incluindo os núcleos locus coeruleus, A5, A1, A2, A7, SubCV e SubCD como esperado (figura 1B). Para confirmar que os neurônios NA expressando o receptor hM4D eram sensíveis ao CNO, nós realizamos gravações em neurônios locus coeruleus (LC), onde observamos uma inibição de neurônios disparando sobre a aplicação de banho CNO (n = 3, Figura 1C).
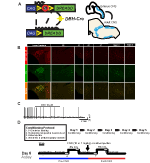
Figura 1. Protocolos esquemáticos e respiratórios de reprodução. O RC::O alelo P_hM4D é combinado com uma linha dopamina-beta-hidroxilase (DBH)-Cre para atingir a expressão específica da célula do receptor hM4D nos neurónios noradrenérgicos (NA) (A). Expressão fluorescente do tdTomato em DBH-Cre; as secções coronais do cérebro do Ai9 Co-expressam-se com a tirosina-hidroxilase (TH) nos núcleos do tronco cerebral (B). Na gravação em fatias de na locus coeruleus antes e depois da aplicação CNO, demonstrando a funcionalidade do receptor DREADD em neurônios-alvo (C). Protocolo respiratório. Os ratos habituados são submetidos a um protocolo habituado de 5 dias que consiste na manipulação, medição da temperatura da sonda rectal, injecção salina e exposição à câmara de pletismografia. No sexto dia, os ratos habituados e não habituados a um ensaio hipercapnico em que o animal é colocado na câmara respiratória e autorizado a aclimatar-se nas condições iniciais de ar ambiente. O animal é então exposto a 20 minutos de 5% de CO2 seguido de 20 minutos de ar ambiente. O animal é então injectado intraperitonealmente com CNO ou solução salina, seguido por mais 20 minutos de ar ambiente pós-injecção, 20 minutos de 5% de CO2, e 20 minutos de ar ambiente. As caixas abertas delineiam os tempos de recolha de dados (D).
CNO-hM4D mediou a perturbação dos neurónios noradrenérgicos em ratinhos adultos
para examinar o papel dos neurónios NA sob a linha de base e a respiração hipercapnica, empregamos o RCC:sistema inibidor P_HM4D cruzado com o driver DBH-Cre. Usando pletismografia de corpo inteiro (Ray et al., 2011), medimos as respostas ventilatórias de animais adultos não arrestados sob o ar ambiente (21% O2/79% N2) e hipercapnic (5% CO2/21% O2/74% N2) condições antes e depois da administração do CNO (figura 1D). Para resolver CNO de dosagem e de potencial tensão induzida pelo nosso projeto experimental, os animais foram submetidos a uma de quatro condições: (1) unhabituated e injetado com 10 mg/kg CNO; (2) habituados e injetado com 10 mg/kg CNO; (3) unhabituated e injetado com 1 mg/kg CNO; ou (4) se habituado e injetado com 1 mg/kg CNO. A habituação consistiu num processo de 5 dias que implicou manuseamento, sonda de temperatura rectal, injecção salina e exposição à câmara de pletismografia todos os dias durante 30 minutos, enquanto os animais naïve só foram expostos a um período de aclimatação da câmara de 20 a 45 minutos imediatamente antes da recolha de dados, como foi feito em estudos anteriores (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Os parâmetros respiratórios medidos incluíram taxa respiratória RR, volume de maré (VT), ventilação mínima (VE), consumo de oxigénio (VO2), ventilação mínima normalizada ao consumo de oxigénio (VE/VO2), frequência de apneia, frequência de sigh e coeficientes de variação para intervalo interbreath e amplitude (instabilidade periódica e volume). Como um controle adicional, nós também comparamos animais de tipo selvagem habitados e não sabituados injetados com solução salina.
CNO-hM4D Perturbation of DBH-Cre Neurons Results in a Hypercapnic Deficit
Under three of the conditions, DBH-Cre; RC::Os animais P_hM4D apresentaram uma redução de VE e VE/VO2 após a administração de CNO, enquanto os controlos irmãos não mostraram diferenças nos valores pré e pós – CNO. Unhabituated animais injetados com 10 mg/kg de CNO mostraram uma reduzida RR (-12.17%, p = 0,034) e VT (-30.87%, p = 0,0016), resultando em uma redução de CINCO (-38.64%, p = 0.0031) e uma ligeira redução no VO2 (-14.25%, p = 0,042) (Figura 2). A redução no VE foi maior do que a diminuição no VO2, resultando em uma redução global no VE/VO2 (-26,89%, p = 0,0095). Unhabituated animais injetados com 1 mg/kg de CNO mostraram uma tendência de redução RR (-12.88%, p = 0.066) e uma diminuição significativa do VT (-16.52%, p = 0.00085) e CINCO (-28.08%, p = 0.0070), levando a uma redução geral na VE/VO2 (-22.23%, p = 0,016) (Figura 3). Finalmente, habituados animais injetados com 1 mg/kg de CNO mostrou uma redução RR (-10.77%, p = 0.074), tendência para a redução de CINCO (-25.07%, p = 0.074), e redução global do VE/VO2 (-23.70%, p = 0,024) (Figura 4). Não foram observadas alterações significativas na frequência da apneia, frequência de suspiro, instabilidade periódica ou volume, ou temperatura em qualquer coorte.
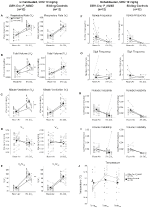
Figura 2. DBH-Cre não embalado; RC:: p_hm4d animais aos quais foi administrado 10 mg/kg de CNO apresentam um défice hipercapnico, enquanto os controlos dos irmãos não mostram qualquer alteração. Após administração do CNO, DBH-Cre; RC::Os animais P_hM4D não apresentam alterações na ventilação do ar ambiente e reduções na RR (a), VT (B), VE (C), VO2 (D) E VE/VO2 (e) em condições hipercapnicas, sem alterações na frequência da apneia (F), frequência de sigh (G), instabilidade periódica (H) ou volume (I), e temperatura (J). Os controlos com irmãos não mostraram diferença em todos os valores pré e pós-CNO.
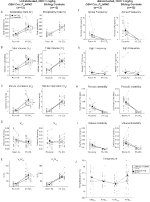
Figura 3. DBH-Cre não embalado; RC:: p_hm4d animais aos quais foi administrado 1 mg/kg de CNO apresentam um défice hipercapnico, enquanto os controlos dos irmãos não apresentam alterações. Após administração do CNO, DBH-Cre; RC::P_hM4D animais mostram nenhuma mudança significativa na sala de ventilação de ar ou RR (UM) e mostrou reduções no VT (B), VE (C), e VE/VO2 (E) em hypercapnic condições, sem alterações no VO2 (D), apnéia frequência (F), suspiro de frequência (G), periódicos (H) ou volume de instabilidade (I), e temperatura (J). Os controlos com irmãos não mostraram diferença em todos os valores pré e pós-CNO.
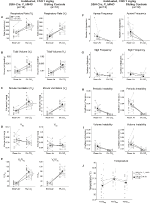
Figura 4. DBH-Cre habitado; RC:: p_hm4d animais aos quais foi administrado 1 mg/kg de CNO apresentam um défice hipercapnico, enquanto os controlos dos irmãos não apresentam alterações. Após administração do CNO, DBH-Cre; RC:: os animais P_hM4D não apresentam alterações na ventilação do ar ambiente e uma redução na RR (a) E VE/VO2 (e) em condições hipercapnicas sem alterações significativas na VT (B), VE (C), VO2 (D), frequência apnea (F), frequência sigh (G), instabilidade periódica (H) ou volume (I) e temperatura (J). Os controlos com irmãos não mostraram diferença em todos os valores pré e pós-CNO.
controlos habitados de irmãos injectados com 10 mg / kg de CNO revelaram um défice ventilatório Hipercapnico
em ambos os DBH-Cre; RC::P_hM4D e irmão de controle habituados animais injetados com 10 mg/kg de CNO, observamos uma redução significativa no VE/VO2 (p = 0.0235) mediada pela diminuição da RR (p = 0.00036) e VE (p = 0,037) (Figura 5). No entanto, ao contrário dos outros grupos, não houve diferença entre DAP-Cre; RC::P_hM4D e irmão de controle de animais nestes parâmetros: RR (-12.72 vs. -10.22%, p = 0.6268), CINCO (-24.88 vs. -23.94%, p = 0.4150), ou VE/VO2 (-15.4 vs. -22.55%, p = 0.4643).
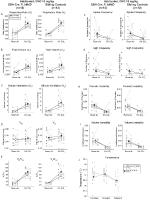
Figura 5. Ambos habitados DBH-Cre; RC::A administração de 10 mg/kg de CNO a animais P_hM4D e controlos de irmãos revela um défice hipercapnico. Depois de CNO administração, DAP-Cre; RC::P_hM4D animais e irmão controles mostram nenhuma mudança na sala de ventilação de ar e uma redução RR (UM), VE (C), e VE/VO2 (E) em hypercapnic condições sem alterações significativas no VT (B), VO2 (D), apnéia frequência (F), suspiro de frequência (G), periódicos (H) ou volume de instabilidade (I), e temperatura (J).
Habituados e Unhabituated tipo selvagem Animais Injetados Com Salina Não Mostrou Alterações Presaline e Postsaline
Apesar de não fenótipos observados em controles injetado com 1 mg/kg de CNO, que abordou a possibilidade de que a injeção em si causou o fenótipo observado nos controles de solo com 10 mg/kg de CNO testando habituados e unhabituated tipo selvagem animais injetados com salina (Figura 6). Tanto nas coortes habitadas como nas Não habitadas, os animais não apresentaram qualquer diferença nos parâmetros respiratórios da administração de presalina e pós-salina. Não foram observadas alterações significativas na frequência da apneia, frequência de suspiro, instabilidade periódica ou volume ou temperatura.
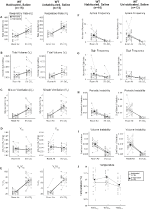
Figura 6. Tanto os animais não vacinados como os animais de tipo selvagem habituados aos quais se administrou solução salina não apresentam diferenças entre os valores pré e pós – injecção. Após a administração salina, tanto os animais do tipo selvagem nãoabituados como os habituados não apresentaram alterações em condições de ar ambiente ou hipercapnicas em RR (a), VT (B), VE (C), VO2 (D), VE/VO2 (e), frequência apnea (F), frequência sigh (G), instabilidade periódica (H) ou volume (I) e temperatura (J).
as razões de concentração da clozapina para CNO são mais elevadas no cérebro do que no soro
para determinar a biodisponibilidade de CNO e clozapina, medimos as suas concentrações no soro e no cérebro através de espectrometria de massa. Trinta minutos após uma injeção intraperitoneal de CNO em ratos, a CNO é encontrado em menor abundância relativa em comparação à sua volta-metabólito clozapina no soro e no cérebro para todas as doses testadas, 0,1 mg/kg (soro p = 0.0054, o cérebro, p = 0,0001) (Figura 7A), 1 mg/kg (soro p > 0.05, o cérebro, p = 0.0197) (Figura 7B), e 10 mg/kg (soro p = 0.0036, o cérebro, p = 0.0005) (figura 7C). Quando geral de clozapina para CNO rácios foram analisados, eles estavam sempre acima de zero e foram mais elevados no cérebro do que no soro do veículo (p > 0.5, 0.1 mg/kg, p > 0,5 a 1 mg/kg, p = 0.0018, 10 mg/kg, p = 0.0160) (Figura 7D).
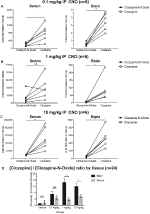
Figura 7. As concentrações de clozapina e CNO são mais elevadas no cérebro do que no soro. Foram colhidas amostras de soro e de cérebro em ratinhos de tipo selvagem 30 minutos após a administração de CNO com 3 doses diferentes de 0, 1 mg/kg (a), 1 mg/kg (B), 10 mg/kg (c). Os pontos de dados individuais representam a abundância relativa de clozapina-N-óxido ou clozapina obtida pelo rácio CLC-MS. clozapina / CNO para cada dose e SEM também são apresentados (D).
discussão
o objectivo inicial deste estudo era examinar o papel dos neurónios DBH-Cre na fisiologia respiratória após perturbação aguda em animais adultos conscientes e desenfreados. Porque os neurônios noradrenalina e NA desempenham um papel bem documentado em comportamentos de estresse (Valentino e Van Bockstaele, 2008; Chen et al., 2018), procurámos também examinar se a habituação a um provável protocolo fisiológico stressante teria um efeito sobre os fenótipos respiratórios previamente observados. Na pletismografia de todo o corpo, o protocolo DREADD usado em nosso laboratório e outros, os animais são manuseados, sondados rectamente para a temperatura, expostos a um ambiente novo (a câmara de pletismografia), e injetados intraperitonealmente. Estudos anteriores mostraram que o manuseio e a habituação à injeção e outros procedimentos de “rotina” podem modificar parâmetros comportamentais e fisiológicos, incluindo respiração (Misslin et al., 1982; Andrews and File, 1993; Lapin, 1995; Ryabinin et al., 1999). Outros estressores aplicados também modificam a respiração sob as condições de base e hipercapnic ventilatory (Isom e Elshowihy, 1982; Kinkead et al., 2001).
nos nossos estudos utilizando doses elevadas de CNO, descobrimos que a inibição mediada pelo hM4D dos neurónios na definidos pelo DBH-Cre resultou numa redução do reflexo hipercapnico no VE/VO2 em quatro grupos experimentais definidos, com redução da RR, VT e VE, apoiando estudos anteriores (Biancardi et al., 2008; Viemari, 2008; Gargaglioni et al., 2010). No entanto, os controlos habitados de irmãos aos quais foi administrada uma dose de 10 mg/kg de CNO utilizada em estudos respiratórios anteriores revelaram um défice ventilatório em condições hipercapnicas da mesma magnitude do observado em DBH-Cre; RC::p_hm4d animais. Nenhum outro grupo de controlo aparentado mostrou este fenótipo, incluindo a coorte habituado que recebeu apenas (1 mg / kg de CNO) ou solução salina. Estes resultados sugerem que doses mais elevadas de CNO (10 mg/kg) têm um efeito na resposta hipercapnica que é desmascarada após uma habituação extensa, ao mesmo tempo que, presumivelmente, reduz os níveis de stress nos animais, e que doses mais baixas de CNO não têm um efeito no controlo respiratório em animais habituados. Estes resultados também estão de acordo com o trabalho de Korsak et al. a OMS demonstrou que uma dose baixa de CNO (2 mg/kg) não produz efeitos fora do alvo na capacidade de trabalho num ensaio de exercício que incluiu formação prévia (Korsak et al., 2018) e Fleury Curado et al. a OMS não mostrou efeitos específicos de dose baixa de CNO (1 mg/kg) na potência respiratória (Fleury Curado et al., 2018).
as razões aumentadas da clozapina para os níveis de CNO no soro e no cérebro (Figura 7) estão em concordância com estudos recentes que sugerem que a CNO é rapidamente metabolizada de volta para clozapina e mostra maior permeabilidade cerebral em comparação com a CNO em ratinhos e em outros lugares (Jann et al., 1994; Chang et al., 1998; Guettier et al., 2009; Gomez et al., 2017; Raper et al., 2017). No entanto, não é claro se os efeitos off-target observados são devidos à CNO ou clozapina. Como as nossas medições hipercapnicas ocorreram < 30 min após a aplicação do CNO, é provável que os efeitos respiratórios fora do alvo sejam mediados pela clozapina. Os nossos resultados (Figura 7) mostram níveis relativos elevados de clozapina no cérebro, embora o CNO não esteja completamente ausente. No entanto, Huckstepp e seus colegas usaram direto CNO aplicativo para o ventral medular em ratos anestesiados para demonstrar que só no ar, e não sob hypercapnic ou hipóxicas desafios, a CNO aplicação tem um pequeno efeito, aumentando a frequência e diminuir expiratória duração, mas deixando VT inalterado, sem claro efeito observado durante hipercapnia (Huckstepp et al., 2015). Dada a aplicação directa em ratos anestesiados e o período de experiência, é provável que os pequenos efeitos fora do alvo observados tenham sido mediados pela CNO e não pela clozapina.
A volta metabólito clozapina é normalmente usado para sedativo e droga antipsicótica, na esquizofrenia, com muitos endógeno, metas, incluindo a baixa-alta afinidade antagônicas ações em D1, D2 e D4 receptores dopaminérgicos, 5-HT2A, 5-HT2C, 5-HT3, 5-HT6 e 5-HT7 serotoninérgicos receptores, os receptores de histamina H1, e α1 e α2 adrenérgicos, entre outros (Fitton e Calcanhar, 1990; Ashby e Wang, 1996). Os efeitos fora do alvo aqui observados podem resultar de mecanismos e metas distintos ou combinados. A clozapina pode afectar a respiração como um sedativo. Um estudo anterior mostrou uma redução de RR e VT abaixo de 5% e 10% de CO2 durante os estados de sono de ondas lentas e de movimento rápido dos olhos, em comparação com o estado silencioso acordado em ratinhos (Nakamura et al., 2007). Alternativamente, a inibição de neurónios que expressam DREADD alvo pode resultar num efeito antianxiogénico ou ansiolítico semelhante ao nosso protocolo de habituação para revelar efeitos quimiossensoriais mediados pela CNO/clozapina fora do alvo. Ambas as explicações são apoiadas por vários estudos que demonstraram que o CO2 desempenha um papel nas respostas inatas e aprendidas ao medo e comportamentos relacionados à ansiedade (Ziemann et al., 2009; Feinstein et al., 2013; Taugher et al., 2014; Dlouhy et al., 2015; Winter et al., 2017). Assim, os neurônios visados em alguns destes estudos podem de fato desempenhar um papel na condução de respostas comportamentais ansiogênicas ao invés de reflexos quimiossensoriais fisiológicos, uma vez que tanto os sistemas catecolaminérgicos quanto serotoninérgicos estão envolvidos em comportamentos de medo/ansiedade e homeostase quimiossensorial (Brust et al., 2014; Hennessy et al., 2017).Por outro lado, os fenótipos quimiossensoriais observados com níveis elevados de CNO podem ser genuínos, pois fomos capazes de recapitular o déficit hipercapnico mediado NA nas doses de CNO, uma magnitude de ordem menor em ratos habitados, enquanto os grupos de controle não mostraram efeito CNO. No entanto, comparações completas em estudos anteriores são difíceis devido à falta, em alguns casos, de dados VO2, VT, RR e VE/VO2 relatados. Por exemplo, as alterações da temperatura do corpo ou taxa metabólica também pode colidir em um número de maneiras sobre o sistema respiratório e chemosensory de saída, e que pletismógrafo câmara temperaturas foram muito diferentes em alguns casos (34 vs. 30°C em nossos estudos), afetando a dinâmica do barométrica componente da forma de onda e, portanto, do volume corrente. Notavelmente, não vimos alterações apreciáveis nas características das formas de onda respiratórias em qualquer uma das nossas condições, nem paragem cardio-respiratória aguda que foi observada nos nossos estudos anteriores sobre o rhombomere em doses elevadas (Sun et al., 2017).
nossos resultados mostram pela primeira vez que o CNO tem um efeito inesperado sobre o reflexo quimiossensorial hipercapnico que é desmascarado pela extensa habituação. Importante, apesar dos altos níveis, um efeito CNO fora do alvo tinha sido previamente descartado devido à falta de fenótipo nos controles de irmãos, mas que mostramos torna-se claro após a habituação. Oferecemos uma caracterização fora do alvo do CNO no sistema modelo do mouse para complementar estudos em ratos e primatas não humanos. Os resultados aqui apontam para a possibilidade de existirem efeitos adicionais mediados pelo CNO, fora do alvo, nos circuitos em estudo ou em circuitos autonômicos ou homeostáticos, mas que podem ser mascarados em outros experimentos controlados. É importante que esses dados revelem que os investigadores devem se esforçar para usar as doses mínimas do ligante ativante possível em combinação com altos níveis de habituação, e que os controles adequados devem ser incluídos nas manipulações genéticas químicas para apreciar e interpretar plenamente os dados experimentais.
materiais e métodos
aprovação ética
estudos foram aprovados pelo Baylor College of Medicine Institutional Animal Care and Use Committee under protocol AN-6171.
reprodução, fundo genético, e manutenção de ratinhos
mantivemos colónias de todas as nossas estirpes heterozigóticas de ratinhos por perfuração posterior a C57BL/6J ratinhos e estirpes homozigóticas por cruzamentos irmãos. Para experimentos de histologia, DBH-Cremice foi acasalado com o rato AI9 homozigous (Madisen et al., 2010) (JAX 007909). Para experimentos de pletismografia, ratos DBH-Cre foram acasalados com RC homozigous:: P_hM4D (Ray et al., 2011) mice to derive animals, in which all mice carried the RC::P_hM4D allele. Os animais irmãos que não herdaram o alelo Cre foram utilizados como animais de controlo para a descendência Cre positiva. Rosa26 primers específicos para o Ai9, RC:: P_hM4D, e RC:: EPE mice were 5′ – GCACTTGCTCCCAAAGTC, 5′ – GGGGTACTTGCATGATGATGAT, and 5′ – CTTAAGCCTGCCAGAAGA and yield a 495 bp band (targeted) and 330 bp band (wt). Os iniciadores específicos do Cre para todos os drivers do CRE rhombomere eram 5′ – ATCGCATCTCAGCAGCAGCACCATTGCCC e 5′ – GCATTTCTGGGATTGCTTA e renderam uma banda de 550 bp se positiva. Para as experiências LC-MS, ratos C57BL/6J foram obtidos a partir do Center of Comparative Medicine (CCM), Baylor College of Medicine.
Histologia
quatro a oito semanas de idade DBH-Cre; ratinhos adultos de Ai9 foram perfundidos transcardiamente com 0, 1 M de solução salina tampão fosfato (PBS) e depois com 4% de paraformaldeído (PFA) em PBS. Os cérebros foram dissecados e a gota fixada para 2 h em 4% de PFA antes de enxaguar e equilibrar em 20% de sacarose em PBS. Os cérebros foram então seccionados em secções de 30 µm, montados em lâminas, e rotulados com anticorpos imunofluorescentes. Nós manchada durante a noite com anti-tirosina hydroxylase de anticorpos para identificar catecholaminergic neurônios (1:1.000, a Millipore AB152), seguido por 2 h com burro anti-coelho Cy3 secundário (1:500, Jackson 711-165-152) em 0,1% de Triton-X em PBS (PBST) com 5% de burro soro. Imagens foram coletadas em um microscópio epifluorescente vertical Zeiss.
electrofisiologia
fatias cerebrais Horizontais contendo o locus coeruleus (300 µm) foram cortadas com um vibratome (Leica VT 1000S, Leica Microsystems, Buffalo Grove, IL) de DBH-Cre adulto; RR2P; RC::ratos ePe (~1 mês de idade) em fluido cerebrospinal artificial de 4°C (ACSF). As fatias foram submergidas em uma câmara perfurada com ACSF oxigenado (95% O2 e 5% CO2) contendo em mM: 124 NaCl, 2.0 KCl, 1.3 MgSO4, 2.5 CaCl2, 1.2 KH2PO4, 25 NaHCO3 e 10 glicose (2-3 ml/min). Gravações de células inteiras foram realizadas a 30 ° C usando técnicas convencionais de grampo patch-clamp e um amplificador MultiClamp 700B (dispositivos moleculares, Union City, CA). Neurônios GFP-positivos do locus coeruleus foram visualmente identificados e posteriormente fotografados por um vídeo de contraste de interferência diferencial infravermelha no palco de um microscópio vertical (Axioskope FS2, Carl Zeiss, Oberkochen, Alemanha). As pipetas (Resistências 4-6 MU) foram cheias com (em mM) 110 K-gluconato, 10 KCI, 10 HEPES, 10 na2-fosfocreatina, 2 Mg3-ATP e 0, 2 na3-GTP; o pH foi ajustado para 7, 2 e a osmolaridade para 300 mOsm. O potencial de detenção era de -70 mV. O CNO foi aplicado em bath.
Plethysmography
Plethysmography on conscious, unrestrained mice was carried out as described on 6-to 12 week-old adult animals (Ray et al., 2011). Os ratos habituados foram submetidos a um protocolo de habituação de 5 dias com cada dia consistindo em vários minutos de manipulação, temperatura tomada por sonda rectal, injecção intra-peritoneal salina e 30 minutos na câmara de pletismografia. Os ratos foram então testados no prazo de uma semana após o último dia de habituação. Os ratos não vacinados não foram expostos ao manuseamento nem à câmara do pletismógrafo. Todos os ratos foram ingênuos com CNO e utilizados apenas uma vez.
no dia do teste, ratos foram retirados de sua gaiola, pesados, e temperatura rectal foi tomada. Os animais foram então colocados numa câmara de pletismografia de fluxo controlado por temperatura (água revestida a 30 ° C) e autorizados a aclimatar-se durante pelo menos 20 minutos em condições de ar ambiente (21% O2/79% N2). Após a aclimatação e medição sob o ar ambiente, o gás de câmara foi transferido para uma mistura hipercapnica de 5% de CO2/21% de O2/74% de N2 durante 20 minutos. O gás de câmara foi então transferido de volta para o ar do quarto por 20 minutos. Os ratos foram brevemente removidos para medição da temperatura rectal e injecção intra-peritoneal de CNO (Instituto Nacional de síntese química de saúde Mental e Programa de fornecimento de medicamentos) dissolvidos em solução salina (1 ou 0, 1 mg/ml) para uma concentração efectiva de 10 ou 1 mg/kg, respectivamente. O animal foi devolvido à câmara por mais 20 minutos de ar ambiente, 20 minutos de hipercapnia, e 20 minutos de ar ambiente. O animal foi então removido da câmara, e a temperatura rectal foi tomada imediatamente após o término da experiência.
Cromatografia Líquida-espectrometria de massa
vinte e quatro ratos de tipo selvagem, divididos uniformemente por sexo, foram pesados e tratados com 10 mg/kg de CNO, 1 mg/kg de CNO, 0,1 mg/kg de CNO, ou veículo. Trinta minutos após a injecção, as amostras de sangue foram recolhidas por punção Cardíaca e colocadas em Microteuros BD. As amostras foram centrifugadas a 4 ° C a 13,500 rpm em centrifugadoras no topo do banco, e os sobrenadantes foram recolhidos. O soro e o cérebro foram mantidos a -20 ° C até à extracção.
solventes de grau HPLC água, acetonitrilo clorofórmio, metanol e ácido fórmico de grau espectrometria de massa foram obtidos de Sigma-Aldrich (St.Louis, MO). A solução de calibração contendo vários calibrantes numa solução de acetonitrilo, ácido trifluoracético e água foi comprada à Agilent Technologies (Santa Cruz, CA). Os metabolitos e os padrões internos, incluindo o ácido N-acetil aspártico-d3, o triptofano-15N2, o ácido glutâmico-d5, a timina-d4, O ácido giberélico, a trans-zeatina, o ácido jasmonico, o ácido 15N antranílico e a testosterona-d3 foram comprados a Sigma-Aldrich (St.Louis, MO). Microtainer R SST TM was obtained from Becton Dickinson (Franklin Lakes, NJ).
a extracção consistiu em 750 µl de metanol frio:água (4:1) contendo 20 µl de padrões internos enriquecidos que foram adicionados a cada amostra cerebral (50 mg) e controlos de qualidade e depois foram homogeneizados durante 1 min cada. Em seguida, adicionaram-se 750 µl de 100% de acetonitrilo contendo 20 µl de padrões internos enriquecidos para lavar a amostra (100 µl) e os controlos de qualidade e, em seguida, sonicaram-se durante 5 minutos. Todas as amostras foram centrifugadas a 5000 rpm durante 10 minutos a 4 ° C. O sobrenadante resultante foi recolhida, e 20 µl foram injetados no LC-MS.
Todas as amostras foram analisadas utilizando-se 6,490 triplo quadripolares espectrômetro de massa (Agilent Technologies, Santa Clara, CA) acoplada ao sistema HPLC (Agilent Technologies, Santa Clara, CA) por vários reação de monitoramento (MRM). Foram obtidos aproximadamente 8-11 pontos de dados por metabolito detectado. Os metabolitos detectados foram clozapina, CNO e norclozapina (n-desmetil clozapina). O modo ESI positivo foi usado no método. A coluna HPLC era a coluna ACQUITYUPLC C18 (100 Å, 1,8 µm, e 2,1 mm × 100 mm. Milford, MA, EUA) com um caudal de 0, 5 ml/min.
Recolha e Análise de Dados
Pletismografia
Pletismografia alterações de pressão foram medidos utilizando um Validyne DP45 transdutor de pressão diferencial e de referência da câmara e CD15 transportadora demodulador e gravou com LabChartPro em tempo real. As formas de onda foram analisadas offline usando o código LabChartPro e MATLAB para determinar a taxa respiratória( RR), volume de maré (VT) (Ray et al., 2011), minute ventilation (VE), and pattern analysis. Ondas respiratórias foram coletadas offline durante os períodos em que o animal estava em repouso, e as leituras estavam livres de artefatos de movimento. Foram analisados, no mínimo, 1 min de dados cumulativos compilados a partir de vestígios de pelo menos 10 s de comprimento a partir dos últimos 10 minutos de uma dada condição experimental. O consumo de O2 foi determinado comparando a composição do gás entre a calibração em uma câmara vazia e a respiração viva usando um sensor de oxigênio e um analisador de AEI. A temperatura da câmara foi constantemente monitorada usando um MicroThermo 2 e sonda e foi registrada com LabChartPro em tempo real.As medições de Poincaré e a frequência de suspiro e apneia foram determinadas usando 1 min de traços livres de movimento de cada condição de respiração. Os suspiros eram definidos como uma respiração com uma amplitude de pelo menos duas vezes maior do que a respiração normal. Apneas foram definidas como um intervalo interbreath (IBI) pelo menos duas vezes maior que o IBI médio. O coeficiente de variação (CV) do IBI e da amplitude foi também calculado a partir da mesma compilação de 1 min de cada condição de respiração (erro padrão IBI ou amplitude/IBI médio ou amplitude).
Estatísticas
Pletismografia
Resultados (RR, VC, VE, VO2, VE/VO2, o número de apnéias e suspiros, e CVs do IBI e amplitude) para o ar ambiente, e hypercapnic dados foram comparados entre DAP-Cre; RC::P_hM4D coortes e irmão controles usando um linear misto efeitos modelo de regressão com o tipo animal (experimental x controle) e CNO administração (pré vs. pós injeção) como efeitos fixos e IDENTIFICAÇÃO do animal como um efeito aleatório. Os dados relativos à temperatura foram comparados utilizando um modelo de regressão linear de efeitos mistos com o tipo animal (experimental vs. controlo) como efeito fixo. Um p < 0, 05 foi usado para indicar significância estatística, e pontos de dados individuais, média e erro padrão da média são mostrados em todas as tabelas.
Cromatografia Líquida-espectrometria de massa
a área obtida sob o pico para cada amostra foi normalizada pelo controle interno e, em seguida, para o veículo antes da análise estatística foi realizada. O teste t não emparelhado foi utilizado para comparar a abundância relativa de clozapina e CNO em cada tecido por grupo de concentração.Todas as experiências foram aprovadas pelo Institutional Animal Care and Use Committee of Baylor College of Medicine. As experiências obedeceram às normas nacionais relativas ao tratamento e à utilização de animais experimentais estabelecidas pela Associação para a avaliação e acreditação dos cuidados laboratoriais com animais.
Autor Contributions
js, FS-M, MC-M e RR conceberam e desenharam os experimentos. JS, FS-M e PZ realizaram os experimentos e contribuíram para a análise dos dados. JS, FS-M, VM, MC-M e RR escreveram o artigo.
Funding
This study was supported by grants R01HL130249 and R01HL130249-S1 from the National Heart, Lung, and Blood Institute; March of Dimes Basil O’Connor Award; and the McNair Medical Institute.
Declaração de conflito de interesses
os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que poderiam ser interpretadas como um potencial conflito de interesses.
agradecimentos
agradecemos aos Drs. Shaila K. Mani e Benjamin Arenkiel pelas discussões úteis. Agradecemos também à Baylor College of Medicine Proteomics Central facility pela execução da cromatografia líquida-espectrometria de massa.
Amiel, J., Dubreuil, V., Ramanantsoa, N., Fortin, G., Gallego, J., Brunet, J.-F., et al. (2009). PHOX2B in respiratory control: lessons from congenital central hypoventilation syndrome and its mouse models. Respir. Physiol. Neurobiol. 168, 125–132. D. O. I.: 10.1016 / j. resp.2009.03.005
CrossRef Full Text | Google Scholar
Andrews, N., and File, S. E. (1993). A história de manipulação de ratos modifica os efeitos comportamentais das drogas no teste elevado de ansiedade. Euro. J. Pharmacol. 235, 109–112. doi: 10.1016/0014-2999(93)90827-5
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S., e Roth, B. L. (2007). Evoluindo a fechadura para encaixar a chave para criar uma família de receptores acoplados à proteína G potentemente ativados por um ligante inerte. Procedimento. Natl. Acad. Ciência. USA 104, 5163-5168. doi: 10.1073 / pnas.0700293104
PubMed Abstract | CrossRef Full Text | Google Scholar
Ashby, C. R., and Wang, R. Y. (1996). Acção farmacológica da droga antipsicótica atípica clozapina: uma revisão. Synapse 24, 349-394. doi: 10.1002/(SICI)1098-2396(199612)24:4<349::AUXÍLIO-SYN5>3.0.CO;2-D
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Bahia, C. M. C. d. S. e Pereira, J. S. (2015). Apneia obstrutiva do sono e doenças neurodegenerativas: uma relação bidirecional. Dement. Neuropsicol. 9, 9–15. doi: 10.1590 / S1980-57642015DN91000003
PubMed Abstract | CrossRef Full Text | Google Scholar
Biancardi, V., Cícego, K. C., Almeida, M. C., and Gargaglioni, L. H. (2008). Neurónios noradrenérgicos Locus coeruleus e condução de CO2 para a respiração. Pflugers Arch. – Euro. J. Physiol. 455, 1119–1128. doi: 10.1007/s00424-007-0338-8
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Brust, R. D., Corcoran, A. E., Richerson, G. B., Nattie, E., e Dymecki, S. M. (2014). Identificação funcional e de desenvolvimento de um subtipo molecular de neurônio serotoninérgico cerebral especializado para regular a dinâmica respiratória. Cell Rep. 9, 2152-2165. doi: 10.1016 / j. celrep.2014.11.027
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Chang, W. H., Lin, S. K., Lane, H. Y., Wei, F. C., Hu, W. H., Lam, Y. W., et al. (1998). Metabolismo reversível da clozapina e clozapina N-óxido em doentes esquizofrénicos. Programa. Neuro-Psicofarmacol. Biol. Psiquiatria 22, 723-739. doi: 10.1016/S0278-5846(98)00035-9
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Chen, Y. W., Das, M., Oyarzabal, E. A., Cheng, P., Plummer, N. W., Silva, K. G., et al. (2018). Identificação genética de uma população de neurónios noradrenérgicos implicada na atenuação das respostas relacionadas com o stress. Mol. Psiquiatria 1-16. doi: 10.1038/s41380-018-0245-8
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kawasaki, H., Oya, H., Buzza, C., et al. (2015). A respiração inibiu quando as convulsões se espalharam para a amígdala e na estimulação da amígdala. J. Neurosci. 35, 10281–10289. doi: 10.1523/JNEUROSCI.0888-15.2015
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Feinstein, J. S., Buzza, C., Hurlemann, R., Follmer, R. L., Dahdaleh, N. S., Coryell, W. H., et al. (2013). Medo e pânico em humanos com danos na amígdala bilateral. Conversao. Neuroci. 16, 270–272. doi: 10.1038 / nn.3323
PubMed Abstract / CrossRef Full Text | Google Scholar
Fitton, A., and Heel, R. C. (1990). Clozapina. Uma revisão de suas propriedades farmacológicas e uso terapêutico na esquizofrenia. Drogas 40, 722-747. doi: 10.2165/00003495-199040050-00007
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Fleury Curado, T. A., Pho, H., Dergacheva, O., Berger, S., Lee, R., Freire, C., et al. (2018). Silenciar os motoneuros hipoglossais leva a uma respiração desordenada em ratos magros. Frente. Neurol. 9:962. doi: 10.3389 / fneur.2018.00962
CrossRef Full Text | Google Scholar
Gargaglioni, L. H., Hartzler, L. K., and Putnam, R. W. (2010). O locus coeruleus e a quimiossensibilidade central. Respir. Physiol. Neurobiol. 173, 264–273. D. O. I.: 10.1016 / j. resp.2010.04.024
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Gomez, J. L., Bonaventura, J., Lesniak, W., Mathews, W. B., Sysa-Shah, P., Rodrigues, L. A., et al. (2017). A quimogenética revelou: ocupação e activação das DREADD através da clozapina convertida. Science 357, 503-507. doi: 10.1126 / science.Aan2475
PubMed Abstract / CrossRef Full Text | Google Scholar
Guettier, J. M., Gautam, D., Scarselli, M., Ruiz de Azua, I., Li, J. H., Rosemond, E., et al. (2009). Uma abordagem química-genética para o estudo G regulação da proteína da função das células beta in vivo. Procedimento. Natl. Acad. Ciência. USA 106, 19197-19202. doi: 10.1073 / pnas.0906593106
CrossRef Full Text | Google Scholar
Hakim, F., Gozal, D., and Kheirandish-Gozal, L. (2012). Alterações simpáticas e catecolaminérgicas na apneia do sono, com especial ênfase nas crianças. Frente. Neurol. 3:7. doi: 10.3389 / fneur.2012.00007
PubMed Abstract / CrossRef Full Text | Google Scholar
Hennessy, M. L., Corcoran, A. E., Brust, R. D., Chang, Y., Nattie, E. E., and Dymecki, S. M. (2017). A actividade dos neurónios Pet1 raphe que expressam a Taquiquininina modula o quimioreflex respiratório. J. Neurosci. 37, 1807–1819. doi: 10.1523/JNEUROSCI.2316-16. 2016
PubMed Abstract | CrossRef Full Text | Google Scholar
Huckstepp, R. T. R., Cardoza, K. P., Henderson, L. E., and Feldman, J. L. (2015). Papel dos núcleos parafaciais no controlo da respiração em ratos adultos. J. Neurosci. 35, 1052–1067. doi: 10.1523/JNEUROSCI.2953-14. 2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Ilg, A.-K., Enkel, T., Bartsch, D., and Bähner, F. (2018). Efeitos comportamentais da clozapina sistémica aguda em ratos de tipo selvagem: implicações para o uso de Rastas na neurociência comportamental. Frente. Behav. Neuroci. 12:173. doi: 10.3389 / fnbeh.2018.00173
CrossRef Full Text | Google Scholar
Isom, G. E., and Elshowihy, R. M. (1982). Interacção do stress agudo e crónico com a respiração: modificação por naloxona. Farmacol. Bioquímica. Behav. 16, 599–603. doi: 10.1016/0091-3057(82)90422-1
PubMed Abstract | CrossRef texto completo | Google Scholar
Jann, M. W., Lam, Y. W., and Chang, W. H. (1994). Formação rápida de clozapina em cobaias e no homem após administração de clozapina-N-óxido. Arco. T. Farmacodyn. Ther. 328, 243–250.
PubMed Abstract | Google Scholar
Joober, R. (2010). Clozapina: uma molécula distinta, pouco compreendida e pouco utilizada. J. Psychiatry Neurosci. 35, 147–149. doi: 10.1503 / jpn.100055
PubMed Abstract / CrossRef Full Text | Google Scholar
Kinkead, R., Dupenloup, L., Valois, N., and Gulemetova, R. (2001). Atenuação induzida pelo Stress da resposta respiratória hipercapnica em ratos acordados. J. Appl. Physiol. 90, 1729–1735. doi: 10.1152/jappl.2001.90.5.1729
PubMed Abstract | CrossRef Full Text | Google Scholar
Klein, D. F. (1993). Falsos alarmes de asfixia, pânico espontâneo e condições relacionadas. Uma hipótese Integrativa. Arco. Gen. Psychiatry 50, 306-317. doi: 10.1001/archpsyc.1993.01820160076009
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Korsak, A., Sheikhbahaei, S., Machhada, A., Gourine, A. V., e Huckstepp, R. T. R. (2018). O papel dos neurônios parafaciais no controle da respiração durante o exercício. Ciência. Rep. 8:400. doi: 10.1038 / s41598-017-17412-Z
CrossRef Full Text | Google Scholar
Lapin, I. P. (1995). Controlos únicos: efeito da manipulação, injeção fictícia e injeção intraperitoneal de solução salina no comportamento dos ratos em um plus-maze elevado. J. Pharmacol. Toxicol. Methods 34, 73-77. doi: 10.1016 / 1056-8719 (95)00025-D
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavezzi, A. M., Alfonsi, G., and Matturri, L. (2013). Patofisiologia do complexo humano de locus coeruleus em morte súbita inexplicável fetal/neonatal. Neurol. Res. 35, 44-53. doi: 10.1179/1743132812Y.0000000108
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
MacLaren, D. A. R., Barreto, R. W., Shaw, J. K., Krishnan Radhakrishnan, S., Khare, P., Espanha, R. A., et al. (2016). Clozapina n-oxide administration produces behavioral effects in long-evans rats: implications for designing DREADD experiments. eNeuro 3, 1-14. doi: 10.1523 / ENEURO.0219-16.2016
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Madisen, L., Zwingman, T. A., Sunkin, S. M., Oh, S. W., Zariwala, H. A., Gu, H., et al. (2010). Um sistema robusto e de alta capacidade de comunicação e caracterização para todo o cérebro do rato. Conversao. Neuroci. 13, 133–140. doi: 10.1038 / nn.2467
PubMed Abstract / CrossRef Full Text | Google Scholar
Mahler, S. V., and Aston-Jones, G. (2018). Não é mau? Considerações para o uso de rastas na neurociência comportamental. Neuropsicofarmacologia 43, 934-936. D. O. I.: 10.1038/npp.2017.299
PubMed Abstract / CrossRef Full Text | Google Scholar
Manvich, D. F., Webster, K. A., Foster, S. L., Farrell, M. S., Ritchie, J. C., Porter, J. H., et al. (2018). O agonista do DREADD clozapina n-oxide (CNO) é metabolizado pelo reverso para clozapina e produz efeitos de estímulo interoceptivo semelhantes à clozapina em ratos e ratinhos. Ciência. Rep. 8, 3840. doi: 10.1038 / s41598-018-22116-Z
CrossRef Full Text | Google Scholar
Misslin, R., Herzog, F., Koch, B., and Ropartz, P. (1982). Efeitos do isolamento, manipulação e novidade na pituitária–resposta adrenal no rato. Psychoneuroendocrinology 7, 217-221. doi: 10.1016/0306-4530(82)90015-4
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Nakamura, A., Zhang, W., Yanagisawa, M., Fukuda, Y., e Kuwaki, T. (2007). Atenuação dependente do Estado de vigilância da quimiorreflex hipercapnica e apneia do sono exagerada em ratinhos knockout orexin. J. Appl. Physiol. 102, 241–248. doi: 10.1152 / japplphysiol.00679.2006
PubMed Abstract / CrossRef Full Text | Google Scholar
Obonai, T., Yasuhara, M., Nakamura, T., and Takashima, S. (1998). Alteração de neurónios de catecolamina no tronco cerebral das vítimas de síndrome de morte súbita infantil. Pediatria 101, 285-288. doi: 10.1542 / Pediatria.101.2.285
PubMed Abstract / CrossRef Full Text | Google Scholar
Ozawa, Y., Takashima, S., and Tada, H. (2003). Alterações do subtipo dos receptores alfa2-adrenérgicos no tronco cerebral na síndrome da Morte Súbita Infantil. Cantarola Cedo. Desenvolvimento. 75 (Suppl.), S129-S138. doi: 10.1016 / j. pathophys.2004. 01. 015
CrossRef Full Text | Google Scholar
Padovan-Hernandez, Y., and Knackstedt, L. A. (2018). Redução Dose-dependente da locomoção induzida pela cocaína pela clozapina-N-óxido em ratos com antecedentes de auto-administração de cocaína. Neuroci. Lett. 674, 132–135. doi: 10.1016 / j. neulet.2018.03.045
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Ramanantsoa, N., Hirsch, M. R., Thoby-Brisson, M.; Dubreuil, V., Bouvier, J., Ruffault, P.-L., et al. (2011). Respiração sem quimiossensibilidade CO(2) em mutações condicionais de Phox2b. J. Neurosci. 31, 12880–12888. doi: 10.1523/JNEUROSCI.1721-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Raper, J., Morrison, R. D., Daniels, J. S., Howell, L., Bachevalier, J., Wichmann, T., et al. (2017). Metabolism and distribution of clozapine-N-oxide: implications for nonhuman primate chemogenetics. ACS Chem. Neurosci. 8, 1570–1576. doi: 10.1021/acschemneuro.7b00079
PubMed Abstract | CrossRef Full Text | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Soriano, L. P., Nattie, E. E., and Dymecki, S. M. (2013). Egr2-os neurónios controlam a resposta respiratória do adulto à hipercapnia. Brain Res. 1511, 115-125. doi: 10.1016 / j. brainres.2012.12.017
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Ray, R. S., Corcoran, A. E., Brust, R. D., Kim, J. C., Richerson, G. B., Nattie, E., et al. (2011). Diminuição do controlo da temperatura respiratória e corporal após inibição serotoninérgica aguda dos neurónios. Science 333, 637-642. doi: 10.1126 / science.1205295
PubMed Abstract | CrossRef Full Text | Google Scholar
Ryabinin, A. E., Wang, Y. M.= = Ligações externas = = Níveis diferentes de imunoreactividade da Fos após manipulação repetida e stress na injecção em duas estirpes de ratinhos. Farmacol. Bioquímica. Behav. 63, 143–151. doi: 10.1016/S0091-3057(98)00239-1
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Snyder, B., Shell, B., Cunningham, J. T., e Cunningham, R. L. (2017). A hipoxia crónica intermitente induz stress oxidativo e inflamação nas regiões cerebrais associadas à neurodegeneração em fase inicial. Physiol. Rep. 5, 1-13. doi: 10.14814 / phy2.13258
PubMed Abstract / CrossRef Full Text | Google Scholar
Sun, J. J., Huang, T.-W., Neul, J. L., and Ray, R. S. (2017). Genes com padrões embrionários do cérebro hindbrain delineiam populações homeostáticas cardio-respiratórias e metabólicas distintas no adulto. Ciência. Rep. 7:9117. doi: 10.1038 / s41598-017-08810-4
CrossRef texto completo | pesquisador do Google
Sun, J. J., and Ray, R. S. (2017). TG (Th-Cre)FI172Gsat (Th-Cre) define neurônios que são necessários para reflexos hipercapnicos e hipóxicos. Biol. Abrir 6, 1200-1208. doi: 10.1242 / bio.026823
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Taugher, R. J., Lu, Y. Wang, Y., Kreple, C. J., Ghobbeh, A., Ventoinha, R., et al. (2014). O núcleo do leito da estria terminalis é fundamental para o comportamento relacionado com a ansiedade provocado pelo CO2 e pela acidose. J. Neurosci. 34, 10247–10255. doi: 10.1523/JNEUROSCI.1680-14. 2014
PubMed Abstract | CrossRef Full Text | Google Scholar
Valentino, R. J., and Van Bockstaele, E. (2008). Regulação convergente da actividade do locus coeruleus como resposta adaptativa ao stress. Euro. J. Farmacol. 583, 194–203. doi: 10.1016 / j. ejphar.2007.11.062
CrossRef Full Text | Google Scholar
Verbraecken, J., and McNicholas, W. T. (2013). Mecânica respiratória e controlo ventilatório na síndrome de sobreposição e hipoventilação da obesidade. Respir. Res. 14:132. doi: 10.1186/1465-9921-14-132
PubMed Abstract | CrossRef texto completo | Google Scholar
Viemari, J.-C. (2008). Modulação noradrenérgica da rede neural respiratória. Respir. Physiol. Neurobiol. 164, 123–130. D. O. I.: 10.1016 / j. resp.2008.06.016
PubMed Abstract / CrossRef Full Text | Google Scholar
Winter, A., ahlbrand, R., Naik, D., and Sah, R. (2017). Sensibilidade diferencial comportamental à inalação de dióxido de carbono (CO2) em ratos. Neurociência 346, 423-433. doi: 10.1016/j. neurociência.2017.01.003
PubMed Resumo | CrossRef Texto Completo | Google acadêmico
Ziemann, A. E., Allen, J. E., Dahdaleh, N. S., Drebot, I. I., Coryell, M. W., Wunsch, A. M., et al. (2009). A amígdala é um quimiossensor que detecta dióxido de carbono e acidose para provocar o comportamento do medo. Cell 139, 1012–1021. doi: 10.1016/j.cell.2009.10.029
PubMed Abstract | CrossRef Full Text | Google Scholar