一般名:citrus leafminerscientific name:Phyllocnistis citrella Stainton(Insecta:Lepidoptera:Gracillariidae:Phyllocnistinae)
citrus leafminer(CLM)、Phyllocnistis citrella Staintonは、柑橘類および関連するRutaceaeおよび関連するRutaceaeの潜在的に深刻な害虫である。いくつかの関連する観賞植物(Beattie1989、clausen1933、Kalshoven1981)。 CLM(図1)は、以前に1914年に米国で傍受されました(ポートは記載されていません)フィリピンからの柑橘類とアタランティア園芸在庫の輸入(Sasscer1915)。 1993年にフロリダ州で発見されたとき、フロリダ州、米国大陸、西半球の新しい記録であった。
図1. 大人の柑橘類のleafminer、Phyllocnistis citrella Stainton。 ライル*J*バス、フロリダ大学による写真。

分布(トップに戻る)
インドのカルカッタ(Stainton1856)から記載された広範なアジア種(Clausen1931、1933、CAB1970)、CLMは東アフリカ—スーダンからイエメン(Badawy1967)、南アジア—サウジアラビアからインド(Fletcher1920)、インドネシア(Kalshoven1981)、香港および中国、フィリピン(sasscer1915)、台湾(Chiu1985、LoおよびChiu1988)、および南日本(clausen1927)に北。 また、ニューギニアやその周辺の太平洋諸島(CAB1970)でも見られる。 それは1940年以前にオーストラリアに導入され、1995年までに大陸に広がっていた(Beattie and Hardy2004)。 CLMは南アフリカと西アフリカの一部でも発生します(CAB、personal communication)。
西半球では、CLMは1993年5月にフロリダ州で、マイアミ・デイド郡の他の地域のホームステッドとブロワード郡とコリアー郡のいくつかの柑橘類の苗床で最初に発見された。 それ以来、それはすべてのフロリダ州の柑橘類の郡に広がっています。 1994年までに、それはアラバマ州、ルイジアナ州、テキサス州に広がっていた(Nagamine And Heu2003)。 1995年までに、シトラスリーフマイナーは、中央アメリカ、メキシコ西部、およびいくつかのカリブ海の島々で発見された(Jones2001)。 2000年、メキシコから南カリフォルニアに到着した(Grafton-Cardwell et al. 2009年にハワイで初めて検出され、2001年にカウアイ島とマウイ島、2002年にモロカイ島とハワイ(ビッグアイランド)に広がった(Nagamine And Heu2003)。
説明(トップに戻る)
CLMの成体(図2)は、4mmの翼幅を持つ微細な蛾です。 彼らは前翅に白と銀色の虹色の鱗があり、いくつかの黒と黄褐色のマーキングがあり、各翼端に黒い斑点があります。 後羽と体は白色で、後羽縁から長い縁鱗が伸びている。 翼を折った休息姿勢では、蛾の外観ははるかに小さい(約2.4mm)。 頭は非常に滑らかで白く、haustellumには基底の鱗がありません。 CLMは、通常は葉の腹側にある蛇行した幼虫の鉱山によって最も容易に検出されます。 幼虫は分(最大3mm)、半透明の緑がかった黄色で、葉の鉱山の内側に位置しています。 蛹は特徴的に葉縁の蛹の細胞にあります。 大人は一般に容易に気づかれるには余りにも分でdiurnallyそして夕方に活動的である。
図2。 ピン留めされた大人の柑橘類のleafminer、Phyllocnistis citrella Stainton、(4mmの翼幅)。 ジェフリー W.Lotz、農業と消費者サービスのフロリダ省、植物産業の部門による写真。

生物学(トップに戻る)
CLMの生物学は、Badawy(1967)、Beattie(1989)、Clausen(1927,1931,1933)、Fletcher(1920)、Kalshoven(1981)、Latif and Yunus(1951)を含む多くの研究者によって報告されている。 CLMの卵は宿主の葉の下側に単独で敷かれる。 卵の黄化は2〜10日以内に起こり、幼虫はすぐに葉に入り、摂食を開始する。 幼虫は若い葉(時には若い芽)に蛇行鉱山を作り、葉のカールと重大な傷害をもたらす。 葉の鉱山は、両方の葉の表面が使用されているときに重い蔓延を除いて、通常、腹側の葉の表面にあります。 通常、葉ごとに一つの葉の鉱山だけが存在しますが、重い蔓延は葉ごとに二つまたは三つの鉱山を持っている可能性があり、大きな葉の九つまでの鉱山は、フロリダ州で発見されています。
同様のリーフマイナーと同様に、幼虫は摂食サイクル中に葉の中で保護されます。 幼虫は4つの幼虫を持ち、発達には5日から20日かかります。 蛹化は、葉のわずかなカールの下で、葉の縁にある特別な蛹の細胞の鉱山内にあります。 蛹の発達には6〜22日かかります。 成虫は夜明け頃に出現し、午前中に活動し、他の活動は夕暮れや夜に発生します。 雌は夜と夜に卵を産む(Badawy1967、Beattie1989)。 CLMは、柑橘類の缶詰を広げるのを助けるかもしれない(Hill1 9 1 8,Ando e t a l. 1985年(昭和55年)、鉱山からの落雷により被害を受けた。
年間の世代はほぼ連続しているようで、日本南部では6人(Clausen1931)、インド北部では9人から13人(Lal1950)、インド南部では10人(Pandey and Pandey1964)である。 開発時間の合計は約13-52日です(Pandey and Pandey1964)。 葉のフラッシングサイクルと気象条件に応じて、13までの数が予想される(Jones2001)。 大人はわずか数日しか生きていません。 フロリダ州では、約3週間ごとに新世代が生産されています。
宿主植物(トップに戻る)
CLMは、その範囲内の柑橘類および関連するムツ科の種に共通している(Kalshoven1981)。
CLMは、オレンジ、レモン、ライム、タンジェリンを含むすべての柑橘類の葉に最も一般的に見られます。
ホストとして記録されている他のRutaceaeは次のとおりです:
- Aegle marmelos(L.)Corr. サーブ インドで(Fletcher1920)
- Atalantia sp. フィリピンでは(Sasscer1915)
- Murraya paniculata(L.)Jack. インドでは(Pruthi and Mani1945)
- Poncirus trifoliata(L.)Raf. インドでは(Clausen1933)
- インドネシアでは様々な在来のRutaceae(Kalshoven1981)
その他の報告されたホストには、:
- インドのJasminum sambac(L.)Aiton(Oleaceae)(Fletcher1920)
- 柑橘類のヤドリギ(Loranthus sp. 1921)
- Pongamia pinnata Pierre(Leguminosae)in India(Margabandhu1933)
- Alseodaphne semecarpifolia Nees(Lauraceae)in India(Latif and Yunus1951)
いくつかの他のホストは、CLMのために報告されているが、幼虫は、これらの互換性のないホスト上で彼らのライフサイクルを完了していません:
- インドのMurraya koenigii L.Sprengel(Rutaceae)(Fletcher1920)
- Jasminum sp. そして、インドのJasminum cinnamomum Kobuski(Oleaceae)(Pruthi and Mani1945)
- Dalbergia sissoo Roxb。 インドのEx DC(Leguminosae)(Latif and Yunus1951)
- Salix sp. (Salicaceae)in India(Pruthi and Mani1945)
- Grewia asiatica L.(Tiliaceae)in India(Latif and Yunus1951)
ダメージ(トップに戻る)
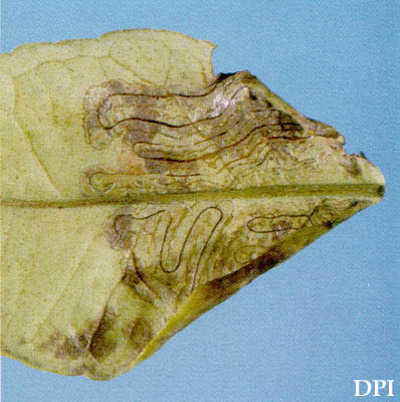
幼虫は葉の中に蛇行鉱山を形成し(図3)、宿主の果実はほとんどありません。 これらの鉱山はfrassの中央線で満たされています。 この特徴は柑橘類のpeelminerからこのleafminerを分けるのを助ける。 柑橘類のleafminerの幼虫はより若い、洗い流す群葉にしか出没しない。 成虫は葉の上面と下面の両方に卵を産み、長さは1.27cm(1/2インチ)未満である。 オーストラリアからの報告は、葉あたり1〜3個の鉱山が蔓延していることを示していますが、フロリダなどの他の地域ではより湿った状態では、葉あたりのより多くの鉱夫をサポートする可能性があります(Jones2001)。
経済的損失は、苗床および非担持柑橘類の保護コストの増加、および戸口栽培者へのホストの販売の制限によって反映されている。 寄生は、若い木の成長を遅らせることができる(Grafton-Cardwell et al. 2009). 重い蔓延は成熟した木の生産に影響を与える可能性がありますが、これはライム(Jones2001)でのみ実証されています。
フロリダ州では、柑橘類のリーフマイナーが、柑橘類のカンカー細菌が木に寄生することを可能にする開口部を作成することが本当の懸念である(Grafton-Cardwell et al. 2009).
(トップに戻る)
:
- ;
- 蛹室は葉縁の近くにあり、その縁はロールオーバーされ、室の露出した部分は明確なオレンジ色であり、
- 緑の芽の多肉植物の枝も攻撃される可能性がある(Battie1989,Pandey and Pandey1964)。
図3。 柑橘類leafminer、Phyllocnistis citrella Stainton、フロリダ州の柑橘類の葉鉱山。 写真:ジェフリー-W. ロッツ、農業と消費者サービスのフロリダ省、植物産業の部門。
管理(トップに戻る)
CLMについて報告された寄生虫には、東南アジア、日本、オーストラリアから39種、主にカルシドイドアが含まれている(Heppner1993,Kalshover1981,Lo and Chiu1988)。 日本では安藤らによってCLMの男性を誘引するフェロモンが開発された。 (7Z,11Z)-7,11-ヘキサデカジエナールと呼ばれる。 フェロモンを含む罠が利用可能であり、蛾がいつ飛行しているかを決定するために使用されるべきである(Grafton-Cardwell et al. 2009)多くの作業は、特にインドでは、化学的制御を使用して行われています。 オーストラリアでは、様々な噴霧レジーム、成長フラッシュの時間、および生物学的制御の促進が推奨されている(Beattie1989)。
フロリダ州では、生物学的制御と油の適用は、CLMの個体数を減らすのに適した方法です。 すでにフロリダ州に存在する天敵は、幼虫と蛹の90%の死亡率を引き起こし、leafminerの蔓延に反応しています。 これらの天敵には、フロリダ州のほとんどに確立され、この死亡率の30%までの責任がある導入された寄生虫Ageniaspis citricolaが含まれています(Rogers、Stansly、Stelinski2009)。 どうやら、Ageniaspis citricolaは柑橘類のleafminerと同時にハワイに導入されました。 それ以来、Ageniaspis citricolaはすべての島に広がり、良いコントロールを提供しています(Nagamine And Heu2003)。
その結果、殺虫剤による治療は寄生虫集団に影響を与える可能性があり、栽培者はそのような治療のタイミングと頻度について州の専門家に相談すべきである(Grafton-Cardwell and Montez2009)。
選択された参照(トップに戻る)
- 安藤T,田口KY,内山M,氏家T,黒子H.1985. (7Z-11Z)-7,11-hexadecadienal性誘引剤,Phyllocnistis citrella Stainton(Lepidoptera,Phyllocnistidae). 農業および生物化学,東京49:3633-3653.
- Badawy A.1967. 形態学とPhyllocnistis citrella Staintの生物学。、スーダンの柑橘類の葉の鉱夫。 エジプト昆虫学会紀要51:95-103.
- 1989. シトラスの葉の鉱夫。 NSW農業&漁業,Agfact,H2. 昭和41年4月に廃止された。
- 1985. 台湾における柑橘類の害虫の生物学的防除。 台湾農業研究所、特別報告書19:1-8。
- 1927. 日本の柑橘類の昆虫。 USDA,Washington,D.C.Technical Bulletin15:1-15.
- 1931. 極東の二つの柑橘類の葉の鉱山労働者。 米国農務省、ワシントンD.C.技術速報252:1-13.
- 1933. 熱帯アジアの柑橘類の昆虫。 USDA,Washington,D.C.Circular266:1-35.
- コモンウェルス農業局(CAB)、コモンウェルス昆虫研究所。 1970. フィロクニスティス-シトレラStnt。 害虫の分布図で。 サー A、地図第274号。 株式会社イースタン-プレス、ロンドン、イギリス。
- 1920. インドの昆虫の生活史。 ミクロレピドプテラ インド農務省の覚書6:1-217。
- ヘプナー JB. 1993. 柑橘類のleafminer、フロリダ(鱗翅目Gracillariidae Phyllecnistinae)のPhyllocnistis citrella。 熱帯鱗翅目4:49-64。
- 1918. ノーザンテリトリーにおける柑橘類の缶詰の歴史(他の場所での発生のメモを含む)。 ノーザンテリトリー-オーストラリア速報18:1-8。
- 1981. インドネシアの作物の害虫。 ジャカルタ:Ichtiar Baru。 .
- Margobandhu V.1933. 北部Circarsのオレンジの害虫。 マドラス農業ジャーナル21:60-68。
- 1950. 昆虫-平野で栽培された果樹の害虫。 アグリック アニム ハスブ ウッタル-プラデーシュ州1:30-45。
- ラティフA、ユヌスCM。 1951. パンジャーブ州の柑橘類の葉の鉱夫の食品植物。 昆虫学研究紀要42:311-316.
- Lo KC,Chiu SC. 1988. 台湾における柑橘類の害虫とその天敵のイラスト。 台中台湾農業研究所。 75頁
- Pandey ND,Pandey YD. 1964. シトラノオトギリソウのバイオノミクスシトラノオトギリソウのバイオノミクスシトラノオトギリソウ。 (鱗翅目:Gracillariidae)。 昆虫学のインドジャーナル26:417-423。
- Pruthi HS,Mani MS.1945. インドの柑橘類の昆虫と鉱山の害虫の知識とそのコントロール。 帝国議会農業研究科学モノグラフ16:1-42。
- 再リンクOA、Groff GW。 1921. カオパン種なしシャムプンメロとその文化。 フィリピン科学ジャーナル19:389-437。
- 1915. 1914年に輸入された養樹園の在庫で収集された重要な害虫。 経済昆虫学のジャーナル8:268-270。
- 1856. インドのマイクロ鱗翅目の三種の説明。 ロンドン昆虫学会の取引(n.s.)3:301-304。
- Stelinski LL,Miller JR,Rogers ME. 2008. 非常に低いフェロモン放出速度で非競争的なメカニズムによって媒介される柑橘類のleafminerの交配破壊。 化学生態学のジャーナル34:1107-1113。
- 1991. ダーウィンの四つの柑橘類の品種を攻撃するPhyllocnistis citrella Stainton(鱗翅目:Phyllocnistidae)に関する注意事項。 オーストラリア昆虫学会誌30:77-78。