広告:
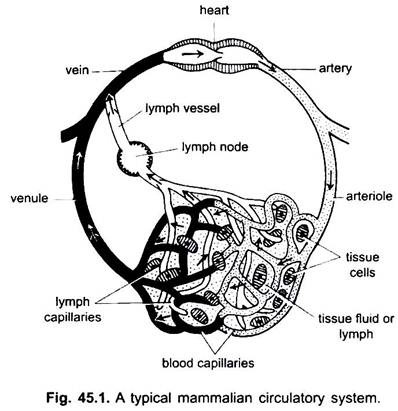
脊椎動物の循環系には、体全体に分岐し、組織に体液を運ぶ精巧に分岐する管の2つのシステムがあります。 それらは血管系およびリンパ系である。 循環系という用語が使用される場合、それは血管系のみを指す。
血管系は脊椎動物の閉鎖系であり、収縮性の心臓と血管と呼ばれる連続した管を持っています。 リンパ系は、リンパ空間を有する開放系である。 心臓から血液を運ぶ血管は動脈であり、これはより薄い細動脈に分割され、非常に薄くて小さな毛細血管に分岐する。
毛細血管の壁は房状の内皮細胞の単層でできています。 心臓を含むすべての血管は、内皮細胞または内皮の内層を有する。 血液は組織細胞と直接接触しません。
:
物質は、細胞間の組織空間に含まれる組織液を通って毛細血管から出入りします。 体の血液細胞と組織細胞との間の物質の交換は、毛細血管壁を介して行われる。 これは、毛細血管の内皮細胞におけるピノサイトーシスによってもたらされる。
電子顕微鏡下では、内皮細胞は原形質膜の陥入である多くの小さな小胞を示す。 これらの小胞は細胞の一方の側から移動し、その内容物を反対側に放出する。
このようにして、血液は代謝のために酸素、栄養物質、ホルモンを細胞に与え、細胞は水、二酸化炭素、窒素性廃棄物を血液中に出して排泄器官に運
毛細血管は軟骨と上皮を除く全ての体組織にネットワークを形成している。 毛管から血は中心の方の血を運ぶ静脈を形作るために結合する薄い細静脈に渡ります。 しかし、いくつかの静脈(門脈、腎静脈、および肝静脈)は、動脈のような毛細血管を有する。 しかし、すべての血液は毛細血管を通って細静脈に通過しません。
:
また、皮膚などのいくつかの臓器にある細動脈と細静脈との間には、いくつかの貫通チャネルがあります。 また、桁に見られる細動脈と細静脈との間の動静脈吻合もある。 そのような接続の機能は明確には分かっていませんが、血圧と循環を調節すると主張されています。
いくつかの場所では、薄い壁の空間または正弦波を介して組織と血液との間の物質交換が行われます。 特定の臓器では、血管はrete mirabile(腎臓、空気膀胱)と呼ばれる小さな血管のコイル状のネットワークを形成することがあります。
循環系の一部:
広告:
脊索動物では、循環系は閉鎖型である。 このタイプは環形動物(無脊椎動物)にも見られます。 別のタイプは、毛細血管が見つからない開放型である。 それは軟体動物および節足動物に見られる。 ここでは、血液は動脈を通って様々な器官に流れ、血液腔または洞を通過し、再び血管(静脈)に入り、心臓に至る。
循環系には、心臓、動脈、静脈、毛細血管、血液が含まれます。 心臓は、筋肉の壁を有する修正された血管であり、定期的に収縮して、明確な血管を介して身体の様々な部分に血液を送り込む。
動脈とその枝は、心臓から血液を運ぶ動脈系を形成します。 静脈およびそれらの支流は、動脈または細動脈の毛細血管から血液を採取し、それを心臓に運ぶ静脈系を構成する。
ポータルシステム:
ポータルシステムでは、血液は心臓に直接戻されませんが、血液が戻る過程で介在する器官(肝臓または腎臓)があります。 血を持って来る静脈は毛管で始まり、毛管、求心性および遠心性の容器として機能する関係した静脈の端動脈のような毛管で、求心性の容器ちょうど終
すべての脊椎動物は、血液が肝臓の毛細血管の二組に通過する肝臓門脈系を持っています。 下部脊椎動物および高等脊椎動物の胚は、血液が心臓に到達する前に腎臓の毛細血管の二組を通過する腎門脈系を有する。 下垂体腺の毛細血管は、小さいが重要なシステムである下垂体門脈システムを形成する。
リンパ系:
環節と軟骨魚類を除く脊索動物に見られる。 リンパ管やリンパ管が含まれます。 リンパは、体細胞の中に見られる組織液です。 それは血漿から赤血球といくつかのタンパク質を引いたものです。
リンパ毛細血管は、リンパを集める薄い盲目の血管のネットワークを形成する。 リンパ管は、毛細血管の結合によって形成される薄肉の血管である。 これらは静脈に空になります。 リンパ節は哺乳動物のリンパ管に見られる。 これらは、疾患に対する身体防御のために使用される血液のリンパ球を形成する。
脊椎動物における心臓の進化:
広告:
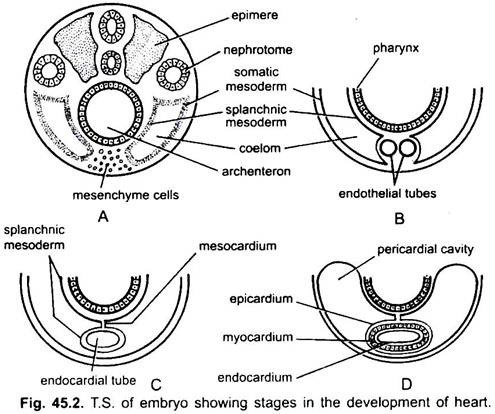
心臓は対になっていない器官であるが、その起源は両側である。 胚では、間充織は咽頭の下に心内膜細胞の群を形成する。 これらの細胞は、一対の薄い内皮管を形成するように配置されるようになる。 二つの内皮管はすぐに咽頭の下に縦方向に横たわっている単一の心内膜管を形成するために融合します。
内胚葉の下にある内胚葉は、心内膜管の周りで縦方向に折り畳まれます。 この二層チューブは、中胚葉が肥厚して心臓の心筋または筋肉壁、および外側の薄い心外膜または内臓心膜を形成する心臓を形成する。 心内膜チューブは、心内膜として知られている心臓のライニングになります。
splanchnic mesodermの折り目は、体腔内の心臓を中断する背側心中膜を形成するために上記を満たしています。 すぐに横方向の中隔が心臓の後ろに形成され、これは体腔を2つの室、心臓を囲む前心膜腔および後腹部腔に分割する。
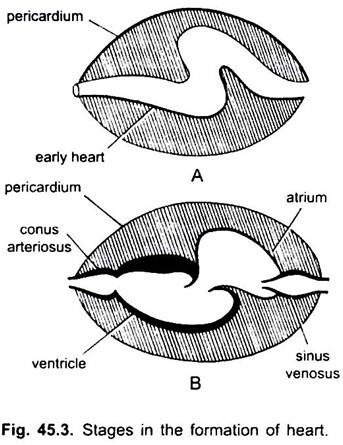
心臓は直管ですが、その端が固定されているため長さが長くなりS字型になります。 弁の出現、狭窄、心臓の仕切り、およびその壁の差動肥厚は、心臓に3つまたは4つの室を形成する。
1. 単房の心臓:
Amphioxus(原始脊索動物)では、真の心臓は発見されていない。 咽頭の下の腹側大動脈の一部は筋肉および収縮性であり、心臓として作用する。
2. 二つのチャンバーハート:
環形動物には、細い壁の静脈洞、わずかに筋肉の心房(耳介)、筋肉の心室、筋肉の動脈瘤または球節の4つの室が線形の順序で配置されています。 それは、他の内臓器官も存在する体腔にある。
4つの部屋のうち、心房と心室のみが高等脊椎動物の4つの部屋(対になった心房と対になった心室)に対応しています。 心臓の進化では、多くの変化が起こっています。
:
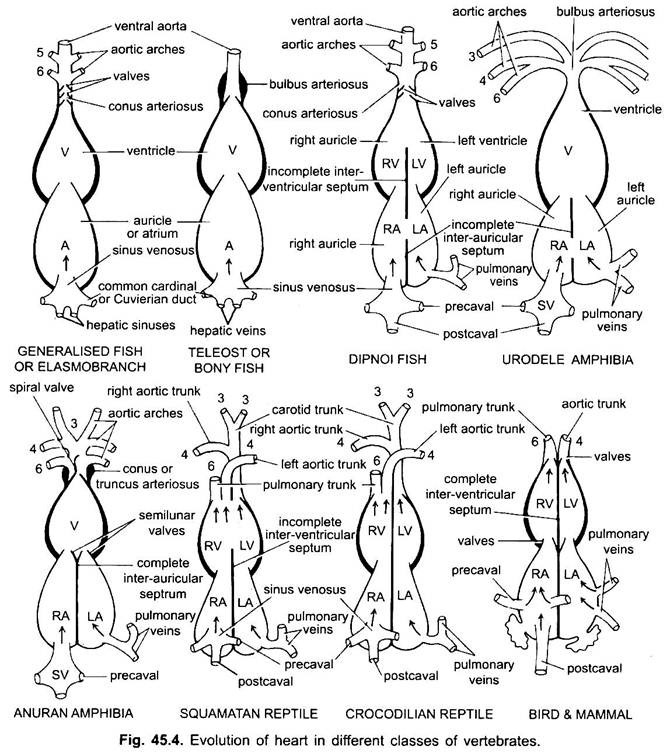
環形動物から硬骨魚類までの魚類の循環系であるDipnoiを除いては、酸素化されていない血液のみが心臓に行き、そこから鰓に送り込まれ、通気され、体に分配される。 軟骨性ドッグフィッシュの心臓は筋肉質で背側に曲がったS字型の管であり、線形の四つの区画がある。
彼らは静脈血を受け取るための洞静脈と心房、そしてこの血液を汲み上げるための心室とconus arteriosusです。 心臓は枝静脈心臓である。 静脈洞および動脈conusは副室である。 心房と心室は真の部屋であり、したがって、それは2室の心臓である。
静脈洞は、一対の弁によって守られた静脈洞-心房開口を介して心房に前方に開く。 心房は心室に背側にあり、一対の弁によって守られた心房-心室開口を介して心室に腹側に開く。 厚い壁の筋肉の心室は、二つのシリーズの弁を含む狭いconus arteriosusに開きます。
心臓は、体腔から横方向の中隔によって分離された心膜腔内に囲まれている。 円錐は心膜を貫通し、腹側大動脈と連続的になる。 心膜腔は、横隔壁の二つの穿孔を介して体腔と連通している。
硬骨魚類:
彼らの心臓はクラスモブランチの心臓に似ています。 硬骨魚類では、円錐は減少し、単一の一対の弁を有する。 Conusに近い腹側大動脈の近位部分は、bulbus arteriosusと呼ばれる、非常に拡大し、厚い壁になります。 それは弾性であり、心室収縮時に拡張する。 心臓は、このように、血液の単一の循環と2チャンバーされています。
3. 三室の心臓:
Dipnoi:
diphoansでは、中隔は心房を左右の室に分けます。 これは、呼吸器官としての水泳膀胱の使用と相関しており、酸素化された血液と酸素化されていない血液の両方が心臓に入り、別々に保たれる二重型循環系の発達に向けた最初のステップを表している。
肺魚の右耳介からの血液は右心室に入り、次に第6対の大動脈弓から分岐する肺動脈によって原始的な肺様ガス膀胱に圧送される。 酸素化された血液は、両生類のような肺静脈を通って左心房に戻る。
両生類:
両生類では、背側心房が心室に前方にシフトする。 静脈洞は右心房に後方に開き、後方には開きません。 心房は完全に右室と左室に分かれており、耳介間中隔に卵円孔がなく、dipnoansでは開いたままである。
心室腔内に深いポケットが発達する。 Conus arteriosusは螺旋弁によって全身血管と肺血管に分かれています。 肺の少ないサンショウウオでは、心房中隔は不完全であり、肺静脈は存在しない。
爬虫類:
爬虫類では、心臓がさらに進んでいます。 心房は常に右室と左室に完全に分離されており、多くの形態では洞静脈が右心房の壁に組み込まれている。 心室はまた、ほとんどの爬虫類では中隔によって部分的に分割されており、ワニとワニでは完全に二部屋になっています。
これは、肺から心臓の左側に来る酸素化された血液が、本質的に身体から右側にある非酸素化された血液から分離されることを意味します。 したがって、ワニでは、2つのタイプの血液は完全に分離され、他の爬虫類ではほぼ完全ですが、循環系の他の部分ではいくつかの混合が発生します。
胚性conus arteriosusは2つの血管の代わりに3つに分割されます:
(i)心室の右側から肺に血液を運ぶ肺弓。
(ii)右第四大動脈弓を介して心室の左側から体に血液を運ぶ右全身大動脈。
(iii)左全身性は右心室から左第四大動脈弓に来る。
左心室からの全身大動脈との接触点では、ワニであっても、二つのタイプの血液の混合があるかもしれないPanizzaeの孔と呼ばれる二つの間の開口部が存在 したがって、爬虫類の心臓は両生類の心臓に対する移行心臓を表しています-2つの完全な耳介と2つの不完全な心室は、左右の全身に血液が少し混
4. 4室の心臓:
鳥類と哺乳類:
鳥類では、心室は完全に2つに分割されているため、心臓は4室(2つの耳介と2つの心室)です。 静脈血と動脈血が完全に分離されています。 全身大動脈は左心室を去り、頭と体に血液を運びます。 肺動脈が右心室を去り、酸素処理のために肺に血液を運ぶ間。
したがって、いかなる場所でも血液の混合がない二重循環が存在する。 静脈洞は完全に右耳介に組み込まれており、二つの前腔と後腔を受け取る。 左耳介は肺静脈を介して酸素化された血液を受け取り、動脈瘤は存在せず、肺大動脈は右心室から生じ、単一の全身大動脈は左心室から生じ、両方ともその基部に弁を有する。
脊椎動物の大動脈弓の変更:
胚動脈:
脊椎動物の胚に心臓が形成されているとき、腹側大動脈と呼ばれる血管が咽頭の下の腹側中央に現れ、すぐ 心臓から生じる腹側大動脈は、咽頭の下を前方に走り、前方に一対の外頸動脈に分割されて頭部に入る。
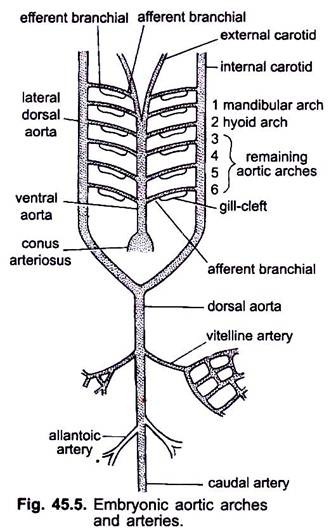
腹側大動脈は、内臓弓を通る6対の外側大動脈弓を等距離で放出する。 それぞれの大動脈弓は、鰓に静脈血を運ぶ腹側求心性分岐動脈と、鰓から酸素化された血液を採取する背側遠心性分岐動脈を有する。 両側の遠心分枝動脈は、内頸動脈として頭部に入る外側背側大動脈または基数大動脈と背側に結合する。
最初の大動脈弓は下顎大動脈弓、2番目の大動脈弓は舌骨大動脈弓、残りの大動脈弓は第3、第4、第5、第6大動脈弓と呼ばれています。 側頭骨大動脈は咽頭の後ろで融合して背側大動脈を形成し、これは尾部動脈として尾部に中背側に続いている。
背側大動脈からは、体の様々な器官を供給する対になった動脈と対になっていない動脈が生じる。 卵黄嚢を有する胚では、一対のビテリン動脈が背側大動脈から生じ、卵黄嚢を供給する。 羊膜の胚では、一対の臍帯動脈または尿膜動脈が背側大動脈から生じ、尿膜に血液を供給する。
成人では、ビテリン動脈が融合して主腸間膜動脈を形成し、尿膜動脈の主要部分は失われるが、それらの残りは下胃動脈または内腸骨動脈を形成す
脊椎動物の大動脈弓:
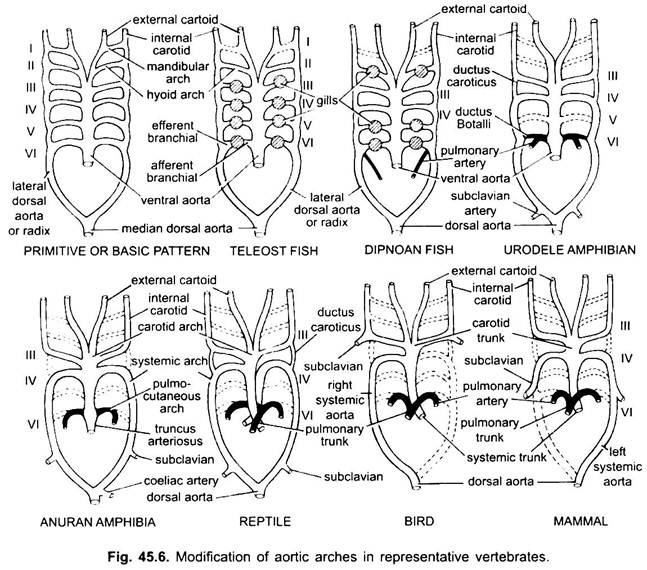
様々な成体脊椎動物では、動脈系は異なっているように見えるが、それらは同じ基本的な基本計画に基づいて構築されている。 違いは、鰓から肺への呼吸の変化による心臓の複雑さの増加によるものです。 脊椎動物シリーズの大動脈弓の数には漸進的な減少がある。
Cyclostomata:
Petromyzonには、大動脈弓の7対があります。 他のサイクロストームでは、これらはMyxineの6対とEptatretusの15対から変化する。
魚座:
魚座の基本的な数であったと考えられているが、魚座の基本的な数であったと考えられている。 この数は、最初のペア、下顎大動脈弓、または遠心性偽頭蓋動脈によって表されるサメおよび光線であっても5に減少する。 ほとんどの骨魚類では、下顎(i)および舌骨(ii)大動脈弓の両方が消失するか、またははるかに減少する。
PolypterusとDipnoi(肺魚)では、えらはよく発達していません。 したがって、肺動脈は、空気膀胱または肺に血液を供給する両側の第6弓の遠心部分から生じる。 ElasmobranchsとDipnoiでは、それぞれの弓は、各鰓に1つの求心性動脈と2つの遠心性枝動脈(分裂によって形成される)を有する。 骨の魚では、各鰓は一つの求心性動脈と一つの遠心性動脈を持っています。
四足歩行大動脈弓では、真の内部鰓が存在しないため、求心性および遠心性の部分に分割されない。 すべてのtetrapodaでは、第一および第二のアーチが消えます。
両生類:
ここでは大動脈弓は鰓の喪失と肺の出現による修正を示しています。 Urodelesには、肺に加えて呼吸器官として存在する外部鰓があります。 III、IV、VおよびVI大動脈弓が存在するが、第五の対はサイレン、AmphiumaおよびNecturusではるかに減少する。 大動脈アーチは、IV、V、およびVI大動脈アーチから生じる枝が外鰓に毛細血管を形成するため、外鰓では求心性および遠心性部分に壊れていない。
III大動脈弓とIV大動脈弓の間の側背大動脈は、血管接続であるcaroticus管として持続する。 VI大動脈弓は肺および皮に血を取る両側のpulmo皮膚の弓か動脈を形作ります。 それはまた動脈管(Botalliの管)として知られている側面背側の大動脈との関係を保ちます。
アヌラン(カエルのオタマジャクシ)の幼虫では、鰓の存在により大動脈弓の配置は成体のurodeleのようである。 変態では、鰓の喪失に伴い、i、II、およびV大動脈アーチは完全に消失し、IIIrd、IVthおよびVlth大動脈アーチのみが存在する。 第三大動脈弓と第四大動脈弓(caroticus管)との間の外側背側大動脈も消失する。 したがって、第三の大動脈弓は、腹側大動脈の一部とともに、酸素化された血液を頭部領域に運ぶ頸動脈弓となる。
側背大動脈に沿った第四大動脈弓は全身弓を形成する。 第六大動脈弓は肺および皮膚に静脈血を供給する肺皮弓となる。 動脈管は変態の間に消失する。 したがって、成体アヌランはIII、IVおよびVI大動脈弓のみを有する。 これらは羊水によっても保持される。
爬虫類:
爬虫類では、鰓は肺に完全に置換されています。 III、IVおよびVI大動脈アーチのみが存在する。 心室を2つの部分に部分的に分離すると、動脈円錐の遠位部分および腹側大動脈全体が3つの血管、すなわち2つの大動脈または全身性および1つの肺に分割される。 右全身弓(IV)は、酸素化された血液を頸動脈弓(III)に運ぶ左心室から生じる。 左全身(IV)および肺大動脈(VI)は、右心室からそれらの起源をとる。 左の全身は、背側大動脈を介して脱酸素化または混合血液を身体に運ぶ。
肺動脈が脱酸素化された血液を肺に取り込む間。 Caroticus管は消えますが、ヘビやいくつかのトカゲ(Uromastix)には持続します。 動脈管はほとんどの爬虫類で消失するが、スフェノドンやいくつかのカメでは減少した形で持続する。 血液の混合のために、爬虫類は魚や両生類のような冷血な動物です。
Aves:
鳥類では、III、IV、VI大動脈弓が存在する。 彼らはいくつかの違いを持つ爬虫類の一般的なパターンに従います。 心室が二つの部分に完全に分割されると、conus arteriosusとventral aortaは二つの血管、左心室から生じる全身大動脈と右心室からの肺大動脈を形成するように分割され
外側大動脈および腹側大動脈の残骸を有する第三大動脈弓は、全身大動脈から生じる頸動脈を形成する。 第四大動脈弓は、右側のみに全身大動脈を形成する。 それはそれ自身の側の側大動脈と結合し、背側大動脈を形成する。 左側の第四大動脈弓の一部は左鎖骨下動脈を形成し、残りはその外側背側大動脈とともに消失する。 第六大動脈弓は肺大動脈を形成する。 Caroticus管と動脈管は消失する。
哺乳類:
哺乳類では、III、IV、VIのaoricアーチも持続する。 心室は完全に2つの部分に分割されています。 <4923><9145>動脈瘤と腹大動脈が分裂して2つの血管を形成する:<4923><9145>(i)左心室から生じる全身大動脈、および<4923><9145>(ii)右心室からの肺大動脈。 側方大動脈および腹側大動脈の残骸を有する第三大動脈弓は、頸動脈弓を形成する。
第四大動脈弓は左側のみで全身大動脈を形成し、右側ではその近位部分は無名および右鎖骨下動脈を形成し、残りは側背大動脈とともに消失する。 第六大動脈弓は肺大動脈を形成する。 動脈管は退化するが、ふ化または出生するまでいくつかの中で細い動脈靭帯として左側の縮小形で持続する。
:
すべての脊椎動物の胚において、静脈系は単純で類似している。 静脈は主に対になって対称的に配置されています。 卵黄嚢のない胚では、腸の下の内臓中胚葉に腸下静脈が形成される。
それは肛門の周りをループし、次のように後方に続けられます:
(i)尾部への尾静脈。 卵黄嚢を有するすべての胚において(卵黄を含むかどうかにかかわらず)、一対のvitelline静脈が卵黄嚢から生じ、静脈洞になる発達中の心臓の後部に結合する。 実際はこれらのvitellineの静脈の融合は骨の多い魚、ハ虫類および鳥の中心の形成に責任があります。 後端の各vitelline静脈は、卵黄嚢を有さない胚と同じように形成された腸下静脈に結合する。
(ii)腎臓の間に一対の脳脊髄下静脈が生じ、尾静脈に合流する。
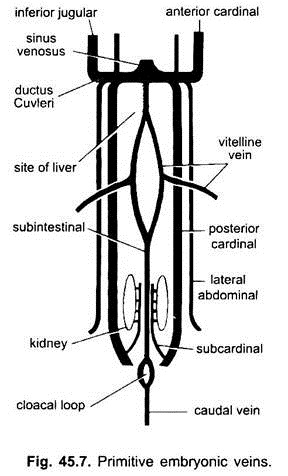
(iii)対の前部および後部の枢機卿静脈が形成され、それぞれ身体の頭部および後部から血液を運ぶ。 各側の前部および後部の枢機卿静脈は洞静脈に入るために横の中隔を通って内部を通る管のCuvieriか共通の枢機卿静脈を形作るために団結します。
魚類およびサンショウウオ(urodeles)では、下頸静脈が頭部の腹側から来て共通の枢機卿静脈に合流する。 それは他の脊椎動物に同族体を持っていません。 羊膜では、一対の側方または腹側の腹部静脈が体壁から来て、共通の枢機卿静脈に入る。
うお座:
共通の枢機卿静脈(キュヴィエの管)は両側から静脈洞に入り、前部枢機卿と後部枢機卿の融合によって形成される。 頭部からの血液は前基数によって収集され、腎臓および生殖腺からの血液は後基数によって収集される。
また、キュヴィエの管に入るのは、体壁から血液を受け取る対の側方腹部静脈と対の付属器です。 腎門脈系は、尾静脈と、腎臓に毛細管現象を起こした腎臓の外側に位置する二つの腎門脈からなる。
肝門脈システムは、胃と腸から血液を運び、肝臓に戻し、一連の正弦波を通過した後、対になったhapatic静脈を介して静脈洞に入る。
硬骨魚類では、外側腹部静脈が欠けているため、胸付属器を排出している鎖骨下からの血液が静脈洞に直接入り、骨盤付属器を排出している腸骨静脈からの血液が後頭骨に入る。
dipnoansでは、単一の腹側腹部静脈が存在し、これはおそらく側腹部の融合に由来する。 この静脈は対になった骨盤の静脈を通ってilacsから血を受け取り、Cuvierの右の管に入る。 なお、右のpostcardinalシステム、新しい静脈から、postclavalは高等脊椎動物の主要な重要性をもつ出現をする。 それは尾静脈と接続され、肝臓を通って洞に前方に通過する。
羊水の胚では、側方腹部静脈は臍帯静脈を排出するため、臍帯静脈または尿膜静脈として知られています。 Vitellineおよび臍帯静脈は出生時に失われ、その残骸のみが持続する。
静脈の修正:
動脈の場合と同様に、異なる脊椎動物の静脈は同じ基本計画に配置されています。 それらによって示されている任意のバリエーションは、脊椎動物シリーズのシーケンスに従います。 その発達において、高等脊椎動物の静脈系は、より低い形態の胚に見られる段階を通過する。
1. 肝臓が形成されているとき、vitelline静脈またはsubtestinal静脈の近位部分は、肝臓と心臓の間の肝静脈を形成し、左vitelline静脈またはsubtestinal静脈の遠位部分は、肝臓に正弦波を形成し、肝臓門脈になり、すべてに存在する肝門脈系を生じさせる。
2. 前枢機卿静脈は内頸静脈として持続する。
3. 魚を除いて、共通の枢機卿静脈は洞静脈(両生類、爬虫類)または右耳介(鳥類、哺乳類)に入る前鼻静脈になる。 鎖骨下静脈は、鎖骨前静脈に合流する各前肢に形成される。
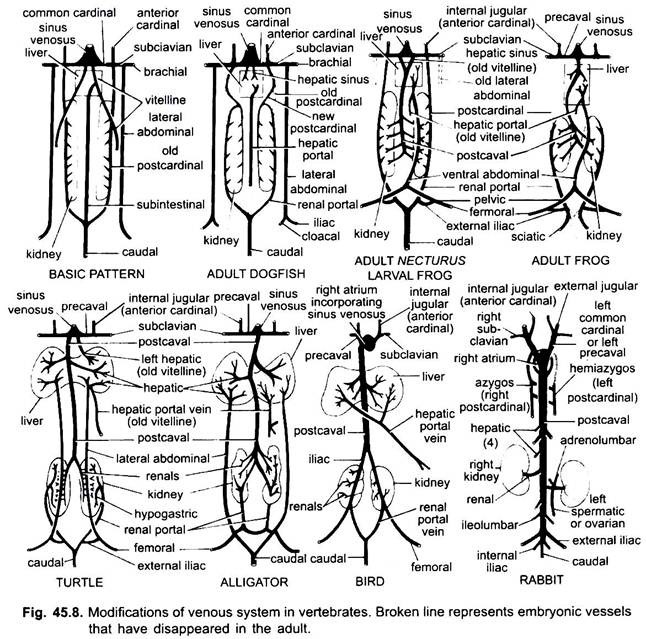
4. 尾静脈は腸下静脈および皮質下静脈との接続を失い、その前部は哺乳類を除くすべての後枢機卿静脈に結合する二つの枝に分割される。
5. 後基静脈は魚類ではそのように持続するが、他のものでは両生類、爬虫類、鳥類では消失する前部の2つの部分に分割されるが、哺乳類では右の前部がアジゴス静脈を形成し、左のものは前鼻との接続を失い、横吻合によってアジゴス静脈に合流するヘミアジゴス静脈を形成することがあり、尾側静脈に合流した後基静脈の後部が腎門脈となる。
各側の腎門脈は腎臓に毛細血管を形成し、魚類、両生類、爬虫類では完全な腎門脈系を形成するが、鳥類では大幅に減少し、哺乳類では後基静脈の後部が消失するため、哺乳類では存在しない。
6. 魚の腎門脈は尾からのみ血液を採取しますが、両生類、爬虫類、鳥類では脚から静脈を奪うこともあります。
7. ある魚およびすべてのtetrapodaでvitellineの静脈はpostcaval静脈を形作るためにsubcardinal静脈を結合します。 両生類および爬虫類では、鼻腔後静脈は腎臓の後端まで伸び、鳥類では腎門脈に結合し、したがって腎門脈系を減少させ、哺乳類では肺後静脈は脚および尾から静脈に結合し、腎門脈系は完全に排除される。
8. 両側腹部静脈は魚ではそのように持続するが、ディプノイから上方に向かって融合して前腹部静脈を形成し、肝臓の近くの肝門脈に合流し、前腹部静脈は腎門脈と肝門脈系を接続する。 腸骨静脈は、前腹部静脈に合流する各後肢に形成される。 前腹静脈は爬虫類でその重要性を失い始め、鳥類では上腹部および尾骨腸間膜静脈を形成するように改変され、哺乳類ではタキグロッサスを除いて消
9. 空気呼吸の形態では、肺静脈は左耳介から成長し、肺に入る。
10. 肺が呼吸器官として機能するようになると、胚の臍帯(allantoic)静脈は消失する。
テトラポダ:
両生類の静脈系は、腹部静脈が静脈洞ではなく肝門系に入ることを除いて、肺魚の静脈系に非常に似ています。 前枢機卿はすべての成体四足動物において内頸静脈として持続する。 下頸静脈は存在しない。 共通の枢機卿は両生類および爬虫類の洞静脈に加わる前大静脈または前大静脈になる。
鳥類および哺乳類では、静脈洞が存在しないため、前腔は心臓の右耳介に直接入る。 人、猫、等で。、左のprecavalは失われます従って左側の血はbrachio頭の枝を通って右のprecavalに入ります。 肺のないサンショウウオでは、心房中隔が不完全であるため、肺静脈は存在しない。
後基静脈は尾静脈を後方に、前基静脈を前方に結合して、幼虫カエルおよびネクトゥルスにおいて共通基静脈を形成する。
爬虫類では、肺静脈および肺後静脈の発達が大きく、身体の後部から腎臓に血液を運ぶ腎門脈システムの重要性が低下しています。
鳥類には、両側に頸椎と鎖骨下の結合によって形成された2つの機能的な前頸静脈と、完全な後頸静脈があります。 それは、腎臓を通過する腎臓ポータルを介して四肢から血液を受け取るが、毛細血管に分割されず、下部脊椎動物の腎臓ポータルに匹敵しない。
哺乳動物では、1つまたは2つの前腔静脈が存在する可能性があります。 発生学的発達が非常に複雑な単一の腹腔後静脈がある。 静脈洞は胚の生活の中で心臓のこの部屋の壁に吸収されるので、すべての大静脈は右の耳介に直接入る。 肝門脈系は他の脊椎動物のそれに非常に似ているが、腎門脈系はない。