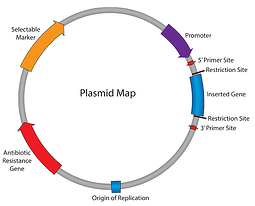
 これまでのところ、私たちのプラスミド101シリーズでは、抗生物質耐性、複製の起源など、プラスミドマップを通して私たちの方法を働いてきました。 この時点まで、我々はプラスミドを複製し、細胞がそれを維持することを確認することができます。 プロモーター-RNAへの挿入物の転写を開始するための責任要素を入力してください。
これまでのところ、私たちのプラスミド101シリーズでは、抗生物質耐性、複製の起源など、プラスミドマップを通して私たちの方法を働いてきました。 この時点まで、我々はプラスミドを複製し、細胞がそれを維持することを確認することができます。 プロモーター-RNAへの挿入物の転写を開始するための責任要素を入力してください。
実際には、用語”プロモーター”は、プロモーター(RNAポリメラーゼ結合部位)と演算子(応答要素)の組み合わせを記載する。 プロモーターは約100〜1000塩基対の長さであり、それらの標的遺伝子の上流に見出される。 プロモーター領域の配列は、RNAポリメラーゼと転写因子の結合を制御するため、プロモーターは、関心のある遺伝子がいつどこで発現されるかを決定する上で大
RNAポリメラーゼ(s)
RNAはRNAポリメラーゼ(RNAP)を使用してDNAから転写されます。 細菌では、これは単一の酵素によって行われる;しかし、真核生物は、それぞれがRnaの特定のサブセットを担当しているmulipleポリメラーゼを持っています。 この特異性を得るために、真核生物のRNAPは、特定のプロモーター要素を認識し、特異的なプロモーター要素に結合することができる。 これは、プラスミド骨格に存在するプロモーターが、作成する必要があるRNAのタイプと互換性がなければならないことを意味します。mRNA(遺伝子発現のために)が必要な場合は、RNAP IIプロモーターを使用する必要がありますが、小さなRna(shRNAなど)はRNAP IIIプロモーターから転写されます。 このポストは、一般的なRNAP IIおよびRNAP III転写のためのプロモーターを備えていますが、rnap IIプロモーターとしてウイルスLTRsを使用することは、一般的にレンチウ
プロモーター特異性
RNA転写産物の種類に基づいてプロモーターを選択する以外に、プラスミドが宿主生物で働くのに適したプロモーターを持っていること 転写機構は細胞の種類や生物によって異なるため、プロモーターは同様に可変でなければならない。 細菌プロモーターは、原核生物細胞においてのみ作用し、典型的には、それらが由来した同じまたは密接に関連する種においてのみ作用する。 同様に、様々な真核細胞型(哺乳類、酵母、植物など)は、ユニークなプロモーターを必要とし、非常に少ないクロスオーバーがあります。 一般的に言えば、細菌のプロモーターは、真核細胞のものよりも少ない部分を有する、あまり多様で複雑ではありません。 他の人がより慎重に制御されている間、いくつかのプロモーターは、恒常的にアクティブであり、すべての時間にあります。 規制されたプロモーターは、特定の組織または開発中の特定の時間にのみ行動するか、化学物質、熱、または光で自由にそれらをオンまたはオフにする方法があるかもしれません。 細胞内では、プロモーター自体は、エンハンサー、境界要素、絶縁体、サイレンサーなどの他の調節因子によって制御されているが、転写のいくつかの”漏れ”が発生する可能性がある。 これは通常、細胞にとって大きな問題ではありませんが、関心のある遺伝子が毒性であれば、研究結果を混乱させたり、細胞を殺すことさえあります。 これに対抗するために、科学者たちは、典型的には他のプロモーター要素のいくつかの組み合わせを含み、より厳密に規制される傾向がある合成プロモーターを作成しました。
真核生物および原核生物の一般的なプロモーター
最も一般的な細菌および哺乳類のプロモーターのいくつかを列挙した以下の2つの参照表を含 これらのリストは決して網羅的ではありませんが、あなたの完璧なプロモーターを選ぶときに始めるのに良い場所でなければなりません!
真核生物プロモーター
| プロモーター | 主に | RNA転写産物 | 説明 | 発現 | 追加の考慮事項 |
| CMV | 一般発現 | mRNA | ヒトサイトメガロウイルス由来の強力な哺乳類発現プロモーター | 構成的 | エンハンサー領域を含むことができます。 いくつかの細胞型で沈黙させることができます。 |
| Ef1A | 一般的な発現 | mRNA | ヒト伸長因子1α | 構成的 | に関係なく、細胞の種類や生理学の一貫した発現を与える傾向があります。 |
| sv4 0<6 7 7 1><1 3 1 8>一般発現<6 7 7 1><1 3 1 8>サル空胞化ウイルス由来のMrna<6 7 7 1><1 3 1 8>哺乳類発現プロモーター4 0<6 7 7 1><1 3 1 8>構成的<6 7 7 1><1 3 1 8>エンハンサー | |||||
| PGK1(ヒトまたはマウス)<6 7 7 1><1 3 1 8>一般発現<6 7 7 1><1 3 1 8>Mrna<6 7 7 1><1 3 1 8>ホスホグリセリン酸キナーゼ遺伝子由来の哺乳動物プロモーター。 | 構成的 | 広範な発現が、細胞型によって異なる場合があります。 メチル化または脱アセチル化によるプロモーターダウン調節に抵抗する傾向がある。 | |||
| Ubc | 一般発現 | mRNA | ヒトユビキチンC遺伝子からの哺乳類のプロモーター | 構成的 | 名前が示すように、このプロモーターは遍在している。 |
| ヒトベータアクチン | 一般発現 | mRNA | ベータアクチン遺伝子からの哺乳類のプロモーター | 構成的 | ユビキタス。 鶏バージョンは、プロモーターハイブリッドで一般的に使用されています。 |
| CAG | 一般発現 | mRNA | 強力なハイブリッド哺乳類プロモーター | 構成 | CMVエンハンサー、鶏ベータアクチンプロモーター、およびウサギベータグロビンスプライス |
| TRE | 一般発現 | mRNA | テトラサイクリン応答要素プロモーター | テトラサイクリンまたはその誘導体で誘導可能。 | は、通常、基礎活性が低い最小限のプロモーターといくつかのテトラサイクリン演算子を含む。 転写は、tet transactivatorが使用されているものに応じてオンまたはオフにすることができます。 |
| UAS | 一般発現 | mRNA | ショウジョウバエプロモーター Gal4結合部位 | 特定の | プロモーターを活性化するためにGal4遺伝子の存在を必要とする。 |
| Ac5 | 一般的な発現 | mRNA | ショウジョウバエのアクチン5c遺伝子 | 構成的 | ショウジョウバエの発現系で一般的に使用される強力な昆虫プロモーター。 |
| ポリヘドリン | 一般発現 | mRNA | バキュロウイルス由来の強力な昆虫プロモーター | 構成的 | 昆虫細胞の発現系で一般的に使用されます。 |
| CaMKIIa | 光遺伝学の遺伝子発現 | mRNA | Ca2+/カルモジュリン依存性プロテインキナーゼIIプロモーター | 特定 | 神経/CNS発現に使用されます。 カルシウムとカルモジュリンで調節されます。 |
| GAL1,10 | 一般発現 | mRNA | 隣接する酵母、多様に転写されたプロモーター | ガラクトースで誘導可能;グルコースで抑制可能 | 独立してまたは一緒に使用 GAL4およびGAL80によって調整される。 |
| TEF1 | 一般発現 | mRNA | 酵母転写伸長因子プロモーター | 構成的 | 哺乳類Ef1Aプロモーターに類似しています。 |
| gds | 一般的な発現 | mRNA | グリセルアルデヒド3-ホスファージデヒドロゲナーゼからの強い酵母発現プロモーター | 構成的 | 非常に強く、TDH3またはGAPDHとも呼ばれる。 |
| ADH1 | 一般的な発現 | mRNA | アルコール脱水素酵素I | エタノールによって抑制される酵母プロモーター | 全長バージョンは、高い発現と強い。 切断されたプロモーターは、より低い発現を有する構成的である。 |
| Camv35S | 一般発現 | カリフラワーモザイクウイルス由来のmRNA | 強力な植物プロモーター | 構成的 | 双子葉植物では活性が高く、単子葉植物では活性が低く、動物細胞では活性がある。 |
| ubi | 一般発現 | トウモロコシユビキチン遺伝子からのmRNA | 植物プロモーター | 構成的 | 植物において高い発現を与える。 |
| H1 | ヒトポリメラーゼIII RNAプロモーターからの小さなRNA発現 | Shrna | 構成的 | は、U6よりもわずかに低い発現を有する可能性がある。 神経細胞でより良い発現を有する可能性がある。 | |
| U6 | ヒトU6小核プロモーター | 構成的 | マウスU6からの小RNA発現 | shRNA | も使用されるが、効率が悪い可能性がある。 |
原核生物プロモーター
| プロモーター | 主に使用される | 説明 | 式 | 追加の考慮事項 | |
| T7 | in vitro転写/一般発現 | T7バクテリオファージからのプロモーター | 構成的であるが、T7RNAポリメラーゼが必要である。 | in vitro転写に使用すると、プロモーターはあなたの遺伝子に対する向きに応じてセンス転写産物またはアンチセンス転写産物のいずれかを駆動します。 | |
| T7Lac | 高レベルの遺伝子発現 | T7バクテリオファージプラスlac演算子からのプロモーター | 誘導されていない場合、基底発現は無視できます。 T7RNAポリメラーゼが必要であり、これはlacオペレーターによっても制御される。 IPTGによって誘導することができます。 | Lac事業者によって非常に厳しく規制されています。 様々な誘導物質濃度を介して遺伝子発現を調節するのに適しています。 | |
| Sp6 | in vitro転写/一般発現 | Sp6バクテリオファージからのプロモーター | 構成的であるが、SP6RNAポリメラーゼが必要である。 | SP6ポリメラーゼは高いプロセス性を有する。 In vitro転写のために使用される場合、プロモーターは、あなたの遺伝子への向きに応じて、センスまたはアンチセンス転写産物のいずれかを駆動します。 | |
| araBAD | 一般的な発現 | アラビノース代謝オペロンのプロモーター | アラビノースとグルコースの存在下で抑制された異化物抑制によって誘導されるか、抗誘 一般にpBADのベクトルで見つけられる。 急速な調節および低い基底の表現のためによい;しかし、いろいろな誘導物質の集中によって調整の遺伝子発現のためにうってつけではない。 | ||
| trp | 高レベルの遺伝子発現 | Eからのプロモーター。 大腸菌のトリプトファンオペロン | 抑制性 | は、高レベルの細胞トリプトファンでオフになります。 | |
| lac | Lacオペロン | からの一般発現プロモーターは、lacリプレッサー(lacIまたはlacIq)の不在において構成的である。 IPTGまたは乳糖によって誘導することができる。 | やや弱い発現を有する漏出性プロモーター。 lacIq変異は、このようにlacプロモーターの調節を締め、リプレッサー10xの発現を増加させます。 様々な誘導物質濃度を介して遺伝子発現を調節するのに適しています。 | ||
| Ptac | 一般発現 | Lacとtrpのハイブリッドプロモーター | Lacプロモーター | のように調節されたlacプロモーター | には、trpBから-35領域とlacから-10領域が含まれています。 非常にタイトな規制。 様々な誘導物質濃度を介して遺伝子発現を調節するのに適しています。 一般的にlac単独よりも優れた表現。 |
| pl | 高レベルの遺伝子発現 | バクテリオファージラムダ | プロモーターは、温度調節可能 | 温度感受性CI857リプレッサーとペアになることが多い。 | |
| T3 | in vitro転写/一般発現 | T3バクテリオファージからのプロモーター | 構成的ですが、T3RNAポリメラーゼを必要とします | in vitro転写に使用すると、プロモーターは遺伝子に対する向きに応じてセンスまたはアンチセンス転写産物を駆動します |
このリストは始めるのに最適な場所ですが、上記の表は、科学者が利用できる組織または開発固有のプロモーターを掘り下げていません。 プラスミドは、多くの場合、治療用途に置かれており、そのような場合には、ここでNIHの研究者によって記載されているように、右の組織特異的プロモーターを同定することが重要です。
注:A.Max Juchheimはこの記事の執筆に貢献しました。