遺伝物質は、染色体と呼ばれる構造の細胞内に貯蔵されており、各末端にテロメアと呼ばれる反復配列を持っています。 特殊なタンパク質はこれらの配列に結合して、染色体を保護し、他の染色体との融合を防止する保護的な「キャップ」を形成する。 酵素のテロメラーゼはまたテロメアの端にDNAの反復的な順序を加えることによって染色体を維持するのを助けます。
最も広く研究されているキャッピング分子の一つは、Cdc13と呼ばれるタンパク質であり、出芽酵母の特定のタイプの一本鎖DNAに結合し、テロメラーゼを 証拠のいくつかの行は、このCST複合体はまた、primase-Polaと呼ばれるDNA複製酵素を募集し、染色体の末端だけでなく、ゲノム内の他の場所でこの酵素の活性を調 2010年、Price et al. とができるようになりました。
同様の複合体は、哺乳類を含む他の真核生物においても同定されており、それらのCst複合体内にStn1、Ten1、およびCTC1と呼ばれる別のタンパク質を含 2010年、Price et al., 2010). しかし、これらのタンパク質の1つだけを欠いている細胞は生き残るのに苦労するため、CST複合体内の異なるタンパク質によって行われる役割を 現在、eLifeでは、Jin-Qiu Zhouと中国科学院とShanghaiTech大学の同僚(最初の著者としてZhi-Jing Wuを含む)が、CST複合体の理解を改善するのに役立つ実験の結果を報告しています(Wu et al., 2020).
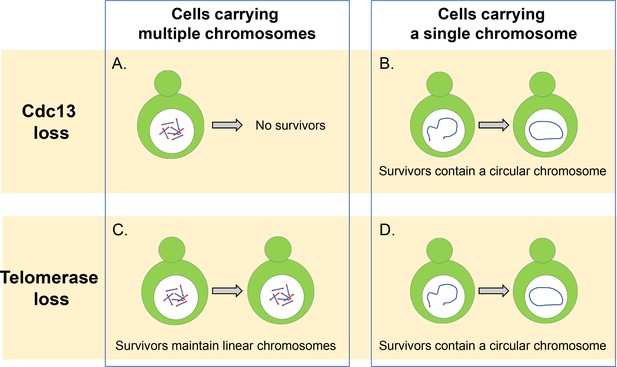
Cdc13とテロメラーゼは、線状染色体を維持するために不可欠です。
(A)複数の線状染色体を有する酵母細胞は、そのテロメアを保護し、染色体が融合するのを防ぐためにキャッピングタンパク質Cdc13を必要とする。 このタンパク質がなければ、これらの細胞は生き残ることができません。 (B)単一の線状染色体を有する細胞は、それらの染色体の末端を一緒に融合させて円形の環を形成することによって、Cdc1 3なしで生存することができ (C)酵素テロメラーゼの非存在下では、複数の線状染色体を有する細胞は、テロメア配列またはクロマチンとテロメア配列の間に位置するDNAセグメントを増幅することができるDNA組換え経路を採用することによって生き残ることができる。 (D)単一の線状染色体を有する細胞は、タンパク質Cdc13を欠いている細胞と同様に、相同組換えを用いて円形染色体を形成するために融合することによ
画像提供:食べログのコンスタンス-ニュージェントさん(元画像)
まず、Wu e t a l. CST複合体を削除すると、その16本の染色体のすべてが一緒に融合して単一の環状染色体を形成した出芽酵母株の生存率にどのように影響するかを調, 2019). 彼らは、円形染色体が反復テロメア配列を含んでいても、CSTを除去しても細胞の増殖を止めたり、より多くの細胞死を招いたりしないことを発見した。 したがって,cst複合体の主な役割は,線状染色体を維持し,染色体が他の染色体と融合するのを防ぐことであり,内部テロメア配列の複製には必須ではないと思われる。
環状の環を形成することに加えて、出芽酵母の16本の染色体を融合させて単一の線状染色体を形成することもできる(Shao et al., 2018). 呉他 CST複合体を除去すると、これらの細胞の生存率が大幅に低下したが、これらの細胞のいくつかは、それらの線状染色体の末端を融合させて円形の環を CST複合体の異なるタンパク質をコードする遺伝子を個別に削除すると、Cdc13を欠いている細胞は、Stn1とTen1の遺伝子を欠いている細胞よりも融合の高 これは、Cdc13が染色体の融合を阻害する上で支配的な役割を果たしていることを示唆しており、Stn1とTen1は独立してCdc13のテロメアの保護に貢献 しかし、このメカニズムの詳細は依然として不明であり、さらなる調査が必要である。
複数の染色体を含む野生型細胞では、テロメラーゼの活性が損なわれていても、融合または環状の染色体を見つけることはまれである。これは、細胞がDNAの二本鎖切断を修復する相同性指向組換えと呼ばれるメカニズムを用いてテロメアを拡張および維持できるためである(図1C)。 しかし、Wu e t a l. 染色体の数を減らすことは、酵素テロメラーゼを欠いている細胞で検出されるより多くの融合につながったことを発見しました。 これは、染色体の数を減らすことは、細胞が環状染色体を産生し、テロメラーゼの損失を生き残ることができる可能性を高めることを示唆している。
特異な線状染色体の両端を融合させることは、非同種末端結合(NHEJ)経路と呼ばれるDNA修復経路に依存すると考えられていた(Haber、2016)。 しかし、Wu e t a l. テロメラーゼの非存在下では、染色体融合は出芽酵母におけるDNA切断の相同組換えに重要な役割を果たしているRad52に依存していることを実証した(図1D)。 単一染色体は染色体の一端近くに逆テロメア配列を有するため、本研究で使用される細胞は染色体循環のためのRad52経路に依存する可能性がある。 このような配列が欠失された場合、細胞は、ヒト細胞においてより一般的なNHEJ経路を介してエンドツーエンド融合を受ける可能性がある(Palm and d e Lange,2 0 0 8)。 追加の実験は、この発見がNHEJ活性の喪失によるものではなく、この経路が出芽酵母細胞において線状化プラスミドを融合させることができることを
呉らの作品。 染色体がどのように融合し、どのようにテロメアがテロメラーゼ酵素から独立して維持されるかについての新しい洞察を提供します。 さらに、この研究からの知見は、酵母を超えて、染色体の末端が融合してリング形状を形成することによって引き起こされる様々なヒト医療症候群の理