PUBBLICITÀ:
Nel sistema circolatorio dei vertebrati ci sono due sistemi di tubi ramificati elaborati, che ramificano in tutto il corpo e trasportano fluidi ai tessuti. Sono un sistema vascolare del sangue e un sistema linfatico. Quando si usa il termine sistema circolatorio, si riferisce solo al sistema vascolare del sangue.
Il sistema vascolare del sangue è un sistema chiuso nei vertebrati; ha un cuore contrattile e tubi continui chiamati vasi. Il sistema linfatico è un sistema aperto con spazi linfatici. I vasi sanguigni che portano via il sangue dal cuore sono arterie, che si dividono in arteriole più sottili, ramificandosi in capillari estremamente sottili e piccoli.
La parete di un capillare è costituita da un singolo strato di cellule endoteliali tassellate. Ogni vaso sanguigno, compreso il cuore, ha un rivestimento di cellule endoteliali o endotelio. Il sangue non entra in contatto direttamente con le cellule dei tessuti.
PUBBLICITÀ:
Le sostanze passano da e nei capillari attraverso il fluido tissutale contenuto negli spazi tissutali tra le cellule. Lo scambio di sostanze tra il sangue e le cellule dei tessuti del corpo avviene attraverso le pareti dei capillari. Ciò è causato dalla pinocitosi nelle cellule endoteliali dei capillari.
Al microscopio elettronico le cellule endoteliali mostrano molte piccole vescicole che sono invaginazioni delle membrane plasmatiche. Queste vescicole si muovono da un lato della cellula e rilasciano il loro contenuto sul lato opposto.
Così, il sangue dà ossigeno, materiale nutritivo e ormoni alle cellule per il metabolismo, e le cellule emettono acqua, anidride carbonica e rifiuti azotati nel sangue per trasportarli agli organi escretori per la loro rapida eliminazione.
I capillari formano una rete in tutti i tessuti del corpo ad eccezione della cartilagine e dell’epitelio. Dai capillari il sangue passa in venule sottili che si combinano per formare vene che portano il sangue verso il cuore. Ma alcune vene (vene portale, vene renali e vena epatica) hanno capillari che sono proprio come quelli delle arterie. Ma tutto il sangue non passa attraverso i capillari nelle venule.
PUBBLICITÀ:
Ci sono anche alcuni attraverso i canali tra arteriole e venule che si trovano in alcuni organi, come la pelle. Ci sono anche anastomosi artero-venose tra arteriole e venule trovate nelle cifre. La funzione di tali connessioni non è chiaramente nota, sebbene si affermi che regolano la pressione sanguigna e la circolazione.
In alcuni punti lo scambio di materiale tra tessuti e sangue avviene attraverso spazi a pareti sottili o sinusoidi. In alcuni organi un vaso sanguigno può formare una rete arrotolata di piccoli vasi sanguigni chiamata rete mirabile (reni, vescica).
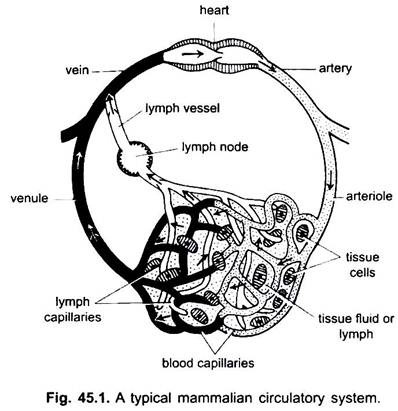
Parti del sistema circolatorio:
PUBBLICITÀ:
Nei cordati, il sistema circolatorio è di tipo chiuso. Questo tipo si trova anche negli anellidi (invertebrati). Un altro tipo è di tipo aperto in cui non si trovano capillari. Si trova nei molluschi e negli artropodi. Qui il sangue scorre attraverso le arterie in vari organi, che passano attraverso gli spazi del sangue o seni e poi di nuovo nei vasi (vene) al cuore.
Il sistema circolatorio comprende cuore, arterie, vene, capillari e sangue. Il cuore è un vaso sanguigno modificato con pareti muscolari, che periodicamente si contrae per pompare il sangue nelle varie parti del corpo attraverso vasi definiti.
Le arterie e i loro rami formano un sistema arterioso che trasporta il sangue dal cuore. Le vene e i loro affluenti costituiscono un sistema venoso che preleva il sangue dai capillari delle arterie o arteriole e lo porta al cuore.
Sistema portale:
Nel sistema portale, il sangue non viene restituito direttamente al cuore, ma c’è un organo interponente (fegato o rene) nel corso del sangue di ritorno. La vena che porta il sangue inizia nei capillari e termina nei capillari, la vena interessata agisce sia come vaso afferente che efferente, i vasi afferenti finiscono nei capillari proprio come le arterie, quindi il sangue viene raccolto nelle vene sistemiche.
Tutti i vertebrati hanno un sistema portale epatico in cui il sangue passa in due serie di capillari nel fegato. Vertebrati inferiori ed embrioni di vertebrati superiori hanno un sistema portale renale anche in cui il sangue passa attraverso due serie di capillari nel rene prima di raggiungere il cuore. I capillari nella ghiandola pituitaria formano un sistema portale ipofisario che è un piccolo ma un sistema importante.
Sistema linfatico:
Si trova nei cordati ad eccezione dei ciclostomi e dei pesci cartilaginei. Include vasi linfatici e linfatici. La linfa è un fluido tissutale che si trova tra le cellule del corpo. È plasma sanguigno meno globuli rossi e alcune proteine.
Capillari linfatici che formano una rete di sottili vasi ciechi che raccolgono la linfa. I vasi linfatici sono vasi a parete sottile formati dall’unione di capillari sanguigni. Questi vuoti nelle vene. I linfonodi si trovano sui vasi linfatici nei mammiferi. Questi formano linfociti di sangue utilizzati per la difesa del corpo contro le malattie.
Evoluzione del cuore nei vertebrati:
PUBBLICITÀ:
Il cuore è un organo spaiato ma la sua origine è bilaterale. In un embrione il mesenchima forma un gruppo di cellule endocardiche sotto la faringe. Queste cellule si dispongono per formare un paio di sottili tubi endoteliali. I due tubi endoteliali si fondono presto per formare un unico tubo endocardico che giace longitudinalmente sotto la faringe.
Il mesoderma splancnico che giace sotto l’endoderma viene piegato longitudinalmente attorno al tubo endocardico. Questo tubo a due strati formerà il cuore in cui il mesoderma splancnico si addensa per formare un miocardio o una parete muscolare del cuore e un epicardio sottile esterno o pericardio viscerale. Il tubo endocardico diventa il rivestimento del cuore noto come endocardio.
Le pieghe del mesoderma splancnico si incontrano sopra per formare un mesocardio dorsale che sospende il cuore nel celoma. Presto si forma un setto trasversale dietro il cuore che divide il celoma in due camere, una cavità pericardica anteriore che racchiude il cuore e una cavità addominale posteriore.
Il cuore è un tubo dritto ma aumenta di lunghezza e diventa a forma di S perché le sue estremità sono fisse. Aspetto di valvole, costrizione, partizioni nel cuore e addensamenti differenziali delle sue pareti formano tre o quattro camere nel cuore.
1. Cuore a camera singola:
In Amphioxus (cordato primitivo), non si trova un vero cuore. Una parte dell’aorta ventrale sotto la faringe è muscolare e contrattile e funge da cuore.
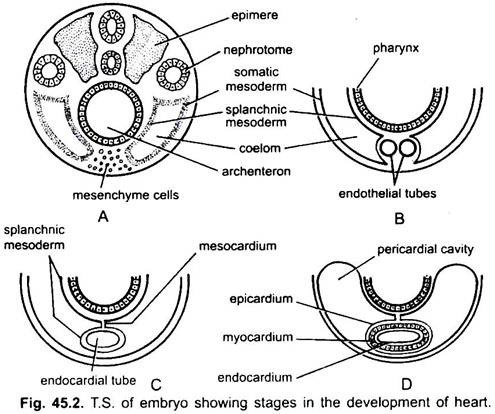
2. Cuore a due camere:
Nei ciclostomi, ci sono quattro camere disposte in ordine lineare: un seno venoso a parete sottile, un atrio leggermente muscolare (padiglione auricolare), un ventricolo muscolare e un conus arteriosus muscolare o bulbus cordis. Si trova nella cavità del corpo in cui sono presenti anche altri organi viscerali.
Su quattro camere, solo atrio e ventricolo corrispondono alle quattro camere (atri accoppiati e ventricoli accoppiati) dei vertebrati superiori. Nell’evoluzione del cuore molti cambiamenti hanno avuto luogo.
Elasmobranchs:
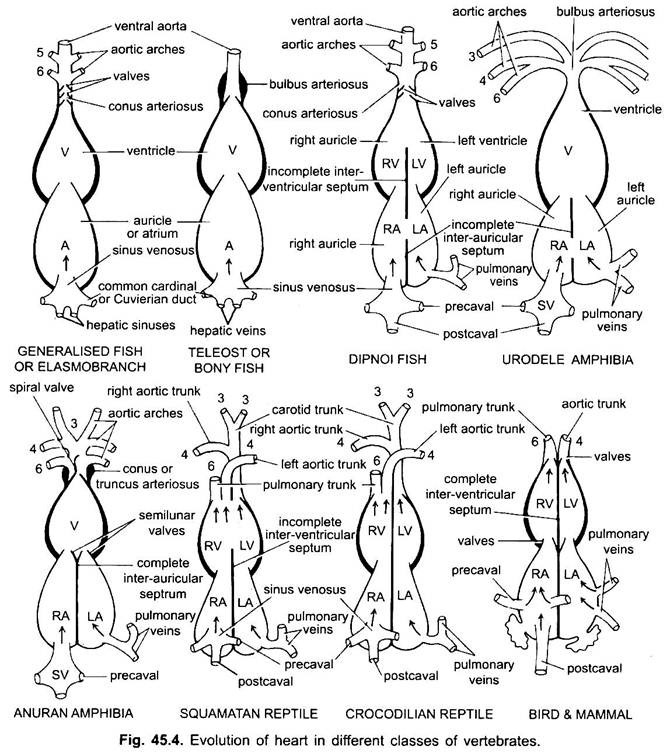
Tranne Dipnoi, il sistema circolatorio nei pesci dai ciclostomi ai teleostei, solo il sangue non ossigenato va al cuore, da lì viene pompato alle branchie, aerato e poi distribuito al corpo. Il cuore di dogfish cartilagineo è muscoloso e dorsoventralmente piegato tubo a forma di S con quattro scomparti in una serie lineare.
Sono il seno venoso e l’atrio per ricevere il sangue venoso e un ventricolo e un cono arterioso per pompare questo sangue. Il cuore è un cuore venoso branchiale. Il seno venoso e il conus arterioso sono camere accessorie. L’atrio e il ventricolo sono vere camere, quindi è un cuore a 2 camere.
Il seno venoso si apre anteriormente nell’atrio attraverso un’apertura sinu-atriale protetta da una coppia di valvole. Atrium giace dorsale al ventricolo e si apre ventralmente nel ventricolo attraverso un’apertura atrio-ventricolare sorvegliata da una coppia di valvole. Il ventricolo muscolare a pareti spesse si apre in uno stretto cono arterioso contenente valvole in due serie.
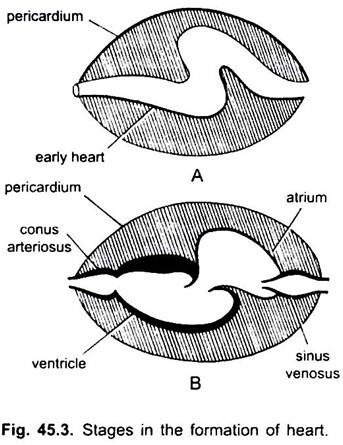
Il cuore è racchiuso all’interno della cavità pericardica separata dalla cavità corporea da un setto trasversale. Conus perfora il pericardio e diventa continuo con l’aorta ventrale. La cavità pericardica comunica con la cavità corporea attraverso due perforazioni nel setto trasversale.
Teleosts:
Il loro cuore assomiglia a quello dei clasmobranchi. Nei teleostati, il cono è ridotto e ha una singola coppia di valvole. La parte prossimale dell’aorta ventrale vicino al conus diventa notevolmente ingrandita e con pareti spesse, chiamata bulbus arteriosus. È elastico e si dilata al momento della contrazione ventricolare. Il cuore è, quindi, a 2 camere con una singola circolazione di sangue.
3. Cuore a tre camere:
Dipnoi:
Nei difoani un setto divide l’atrio in una camera destra e sinistra. Questo è correlato con l’uso della vescica natatoria come organo di respirazione e rappresenta il primo passo verso lo sviluppo del sistema circolatorio a doppio tipo per cui sia il sangue ossigenato che quello non ossigenato entrano nel cuore e vengono tenuti separati.
Il sangue dal padiglione auricolare destro del lungfish passa nel ventricolo destro e viene quindi pompato nella primitiva vescica gassosa simile al polmone dalle arterie polmonari che si diramano dalla sesta coppia di archi aortici. Il sangue ossigenato ritorna nell’atrio sinistro per mezzo di vene polmonari come anfibi.
Anfibi:
Negli anfibi, l’atrio dorsale si sposta anteriormente al ventricolo. Il seno venoso si apre nell’atrio destro dorsalmente e non posteriormente. L’atrio è completamente diviso in camere destra e sinistra e non ha forame ovale nel setto interauricolare, che rimane aperto nei dipnoani.
Tasche profonde si sviluppano nella cavità ventricolare. Il conus arteriosus si divide in vasi sistemici e polmonari da una valvola a spirale. Nei polmoni meno salamandre, il setto interatriale è incompleto e le vene polmonari sono assenti.
Reptilia:
Nei rettili, il cuore è ulteriormente avanzato. L’atrio è sempre completamente separato in una camera destra e sinistra, e in molte forme il seno venoso è incorporato nella parete dell’atrio destro. Il ventricolo è anche parzialmente diviso da un setto nella maggior parte dei rettili, e negli alligatori e nei coccodrilli è completamente a due camere.
Ciò significa che il sangue ossigenato proveniente dai polmoni sul lato sinistro del cuore è essenzialmente separato dal sangue non ossigenato dal corpo sul lato destro. Così, nei coccodrilli, i due tipi di sangue è completamente separato, e quasi completo in altri rettili, ma alcuni miscelazione avviene in altre parti del sistema circolatorio.
Il conus arterioso embrionale si divide in tre invece di due vasi:
(i) Arco polmonare che trasporta il sangue ai polmoni dal lato destro del ventricolo.
(ii) aorta sistemica destra che trasporta il sangue dal lato sinistro del ventricolo al corpo attraverso il quarto arco aortico destro.
(iii) Il sistema sinistro proviene dal ventricolo destro al quarto arco aortico sinistro.
Nel punto di contatto con l’aorta sistemica dal ventricolo sinistro, anche nei coccodrilli, è presente un’apertura tra i due, chiamata forame di Panizzae dove può esserci una certa mescolanza dei due tipi di sangue. Così, cuore rettile rappresenta il cuore di transizione contro cuore anfibio-2 padiglioni auricolari completi e 2 ventricoli incompleti con un po ‘ di miscelazione di sangue in destra e sinistra sistemica.
4. Cuore a quattro camere:
Aves e Mammalia:
Negli uccelli, il ventricolo è completamente diviso in due, in modo che il cuore sia a quattro camere (2 padiglioni auricolari e 2 ventricoli). C’è una completa separazione del sangue venoso e arterioso. L’aorta sistemica lascia il ventricolo sinistro e trasporta il sangue alla testa e al corpo. Mentre l’arteria polmonare lascia il ventricolo destro e trasporta il sangue ai polmoni per l’ossigenazione.
Quindi, c’è una doppia circolazione in cui non c’è miscelazione di sangue in nessun luogo. Il seno venoso è completamente incorporato nel padiglione auricolare destro, che riceve due precavali e un postcavale. Il padiglione auricolare sinistro riceve sangue ossigenato attraverso le vene polmonari, il conus arteriosus è assente, l’aorta polmonare deriva dal ventricolo destro e l’aorta sistemica singola deriva dal ventricolo sinistro ed entrambi hanno valvole alle loro basi.
Modificazioni degli archi aortici nei vertebrati:
Arterie embrionali:
Quando il cuore si forma in un embrione di vertebrati, un vaso sanguigno chiamato aorta ventrale appare a metà ventralmente sotto la faringe, che presto si collega al conus arterioso. L’aorta ventrale derivante dal cuore corre in avanti sotto la faringe e si divide anteriormente in un paio di arterie carotidi esterne in testa.
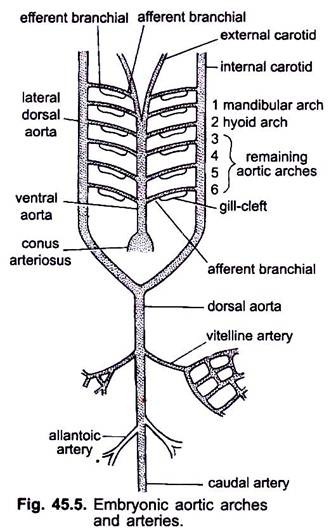
L’aorta ventrale emette 6 coppie di archi aortici laterali a equidistanza che attraversano gli archi viscerali. Ogni arco aortico ha un’arteria branchiale afferente ventrale che trasporta sangue venoso alla branchia e un’arteria branchiale efferente dorsale che preleva sangue ossigenato dalla branchia. Le arterie branchiali efferenti di entrambi i lati si uniscono dorsalmente con l’aorta dorsale laterale o l’aorta radix che entra nella testa come arteria carotide interna.
Il primo arco aortico è un arco aortico mandibolare, il secondo è un arco aortico ioide, i restanti sono chiamati terzo, quarto, quinto e sesto arco aortico. L’aorta dossale laterale si fonde dietro la faringe per formare un’aorta dorsale che viene continuata a metà dorsale nella coda come arteria caudale.
Dall’aorta dorsale sorgono arterie accoppiate e spaiate che forniscono vari organi del corpo. In un embrione con un sacco vitellino, una coppia di arterie vitelline nasce dall’aorta dorsale e fornisce il sacco vitellino. Negli embrioni di amnioti una coppia di arterie ombelicali o allantoiche nasce dall’aorta dorsale che fornisce sangue all’allantois.
In un adulto le arterie vitelline si fondono per formare l’arteria mesenterica principale, la maggior parte delle arterie allantoiche è persa, ma i loro resti formano arterie hypache ipogastriche o interne.
Archi aortici nei vertebrati:
In vari vertebrati adulti, il sistema arterioso sembra essere diverso, ma sono costruiti sullo stesso piano fondamentale di base. La differenza è dovuta alla crescente complessità del cuore a causa di un cambiamento nella respirazione dalle branchie ai polmoni. C’è una progressiva riduzione del numero di archi aortici nella serie dei vertebrati.
Cyclostomata:
In Petromyzon, ci sono 7 coppie di archi aortici. In altri ciclostomi questi variano da 6 coppie in Mixina e 15 coppie in Eptatretus.
Pesci:
Sebbene sei sia considerato il numero base di archi aortici per i pesci. Questo numero è ridotto a cinque anche negli squali e nelle razze con la perdita della prima coppia, l’arco aortico mandibolare o è rappresentato da un’arteria pseudobranchiale efferente. Nella maggior parte dei pesci ossei, gli archi aortici mandibolari (i) e ioidi (ii) scompaiono o sono molto ridotti.
In Polypterus e Dipnoi (polmone-pesci), branchie non sono ben sviluppati. Quindi l’arteria polmonare nasce dalla parte efferente del sesto arco su entrambi i lati che fornisce sangue alla vescica o al polmone. In elasmobranchs e Dipnoi ogni arco ha un’arteria branchiale afferente e due efferenti (formate dalla scissione) in ciascuna branchia. Nei pesci ossei ogni branchia ha un’arteria afferente e un’arteria efferente.
In tetrapoda gli archi aortici non si rompono in parti afferenti ed efferenti perché le vere branchie interne sono assenti. In tutti i tetrapoda il primo e il secondo arco scompaiono.
Amphibia:
Qui gli archi aortici mostrano modifiche dovute alla perdita di branchie e all’aspetto dei polmoni. In urodeles ci sono branchie esterne presenti come organi respiratori oltre ai polmoni. Sono presenti gli archi aortici III, IV, V e VI, anche se la quinta coppia è molto ridotta in Siren, Amphiuma e Necturus. Gli archi aortici non sono rotti nelle branchie esterne in porzioni afferenti ed efferenti, perché i rami derivanti dagli archi aortici IV, V e VI formano capillari nelle branchie esterne.
L’aorta dorsale laterale tra gli archi aortici III e IV persiste come connessione vascolare, il dotto caroticus. L’arco aortico VI forma l’arco o l’arteria pulmo-cutanea su entrambi i lati portando il sangue al polmone e alla pelle. Mantiene anche una connessione con l’aorta dorsale laterale nota come dotto arterioso (dotto di Botalli).
Nella larva di anuran (girino di rana), la disposizione degli archi aortici è come un urodele adulto a causa della presenza di branchie. Alla metamorfosi, con la perdita delle branchie, gli archi aortici I, II e V scompaiono completamente, sono presenti solo gli archi aortici IIIrd, IVth e Vlth. Scompare anche l’aorta dorsale laterale tra il terzo e il quarto arco aortico (dotto carotico). Pertanto, il terzo arco aortico insieme a una parte dell’aorta ventrale diventa l’arco carotideo che trasporta sangue ossigenato nella regione della testa.
Il quarto arco aortico lungo la sua aorta dorsale laterale forma l’arco sistemico. Il sesto arco aortico diventa l’arco pulmocutaneo che fornisce sangue venoso a polmoni e pelle. Il dotto arterioso scompare durante la metamorfosi. Pertanto, gli anuri adulti hanno solo archi aortici III, IV e VI. Questi sono anche trattenuti dagli amnioti.
Reptilia:
Nei rettili, le branchie sono completamente sostituite dai polmoni. Sono presenti solo archi aortici III, IV e VI. Con la separazione parziale del ventricolo in due parti, la porzione distale del cono arterioso e l’intera aorta ventrale sono divise in tre vasi, cioè due aortici o sistemici e uno polmonare. L’arco sistemico destro (IV) deriva dal ventricolo sinistro che trasporta sangue ossigenato all’arco carotideo (III). L’aorta sistemica sinistra (IV) e polmonare (VI) prendono la loro origine dal ventricolo destro. Il sistemico sinistro trasporta sangue deossigenato o misto al corpo attraverso l’aorta dorsale.
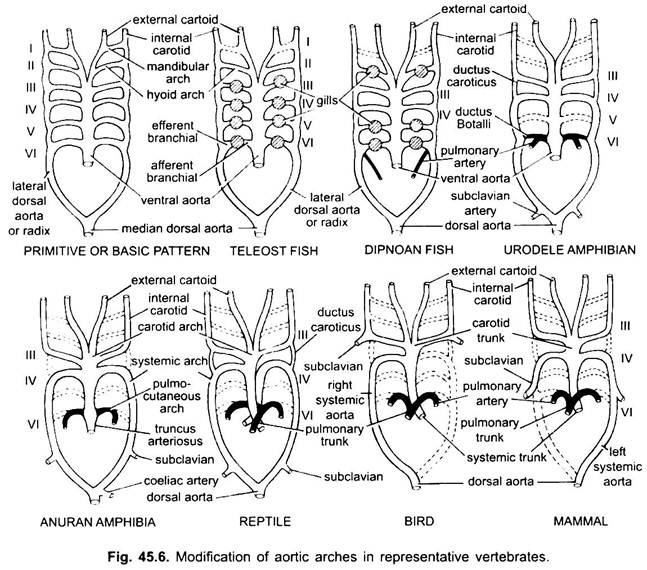
Mentre l’arteria polmonare porta sangue deossigenato ai polmoni. Il dotto caroticus scompare, ma persiste nei serpenti e in alcune lucertole (Uromastix). Il dotto arterioso scompare nella maggior parte dei rettili anche se persiste in forma ridotta in Sphenodon e alcune tartarughe. A causa della miscelazione del sangue, i rettili sono animali a sangue freddo come pesci e anfibi.
Aves:
Negli uccelli sono presenti gli archi aortici III, IV e VI. Seguono il modello generale dei rettili con alcune differenze. Con la divisione completa del ventricolo in due parti, il cono arterioso e l’aorta ventrale si sono divisi per formare due vasi, l’aorta sistemica derivante dal ventricolo sinistro e un’aorta polmonare dal ventricolo destro.
Il terzo arco aortico con resti di aortae laterali e ventrali forma le carotidi che derivano dall’aorta sistemica. Quarto arco aortico forma l’aorta sistemica solo sul lato destro. Si unisce con l’aorta laterale del proprio lato e forma l’aorta dorsale. Parte del quarto arco aortico del lato sinistro forma l’arteria succlavia sinistra, il resto insieme alla sua aorta dorsale laterale scompare. Il sesto arco aortico forma l’aorta polmonare. Il dotto carotico e il dotto arterioso scompaiono.
Mammalia:
Nei mammiferi persistono anche gli archi aorici III, IV e VI. Il ventricolo è diviso completamente in due parti.
Il cono arterioso e l’aorta ventrale si dividono per formare due vasi:
(i) Un’aorta sistemica derivante dal ventricolo sinistro e
(ii) Un’aorta polmonare dal ventricolo destro. Terzo arco aortico con resti di aorta laterale e ventrale forma l’arco carotideo.
Il quarto arco aortico forma l’aorta sistemica solo sul lato sinistro, mentre sul lato destro la sua porzione prossimale forma un’arteria succlavia innominata e destra, il resto insieme alla sua aorta dorsale laterale scompare. Sesto arco aortico forma l’aorta polmonare. Il dotto arterioso degenera ma persiste in alcuni fino alla schiusa o alla nascita in forma ridotta sul lato sinistro come un sottile legamento arterioso.
Sistema venoso:
Vene embrionali:
In tutti gli embrioni vertebrati, il sistema venoso è semplice e simile. Le vene sono per lo più accoppiate e disposte simmetricamente. Negli embrioni senza sacco vitellino, una vena sub-intestinale si forma nel mesoderma splancnico sotto l’intestino.
Si avvolge intorno all’ano e viene continuato posteriormente come:
(i) Una vena caudale nella coda. In tutti gli embrioni che hanno un sacco vitellino (che contenga tuorlo o meno), una coppia di vene vitelline nasce dal sacco vitellino e si unisce alla parte posteriore del cuore in via di sviluppo che diventa il seno venoso. Infatti la fusione di queste vene vitelline è responsabile della formazione del cuore in pesci ossei, rettili e uccelli. Ogni vena vitellina alla sua estremità posteriore si unisce alla vena subintestinale formata allo stesso modo degli embrioni senza sacco vitellino.
(ii) Una coppia di vene subcardinali sorge tra i reni e si unisce alla vena caudale.
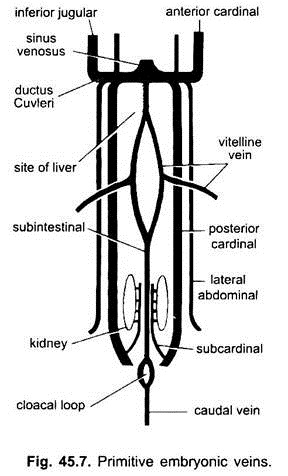
(iii) Si formano vene cardinali anteriori e posteriori accoppiate, che trasportano rispettivamente il sangue dalla testa e dalle parti posteriori del corpo. Le vene cardinali anteriori e posteriori di ciascun lato si uniscono per formare un dotto Cuvieri o vena cardinale comune che passa verso l’interno attraverso il setto trasversale per entrare nel seno venoso.
Nei pesci e nelle salamandre (urodeles) una vena giugulare inferiore proviene dal lato ventrale della testa per unirsi alla vena cardinale comune. Non ha omologhi in altri vertebrati. In un amniotes un paio di vene addominali laterali o ventrali proviene dalla parete del corpo per entrare nelle vene cardinali comuni.
Pesci:
La vena cardinale comune (dotto di Cuvier) entra nel seno venoso da ciascun lato ed è formata dalla fusione di cardinali anteriori e posteriori. Il sangue dalla testa viene raccolto dai cardinali anteriori e il sangue dai reni e dalle gonadi viene raccolto dai cardinali posteriori.
Anche entrando nei condotti di Cuvier sono le vene addominali laterali accoppiate che ricevono sangue dalla parete del corpo e appendici accoppiate. Il sistema portale renale è costituito dalla vena caudale e da due vene portale renali situate lateralmente ai reni che si sono capillarizzate nei reni.
Il sistema portale epatico trasporta il sangue dallo stomaco e dall’intestino e lo restituisce al fegato, dove, dopo aver attraversato serie di sinusoidi, entra nel seno venoso per mezzo di vene apatiche accoppiate.
Nei teleostei, le vene addominali laterali sono carenti in modo che il sangue dai succlavi, drenando le appendici pettorali, entri direttamente nel seno venoso, e il sangue dalle vene iliache, drenando le appendici pelviche, passa nei postcardinali.
Nei dipnoani, è presente una singola vena addominale ventrale, presumibilmente derivata da una fusione di addominali laterali. Questa vena riceve sangue dagli ilac per mezzo di vene pelviche accoppiate ed entra nel dotto destro di Cuvier. Inoltre, dal sistema postcardinale destro, una nuova vena, il postclavale fa la sua apparizione che è di grande importanza nei vertebrati superiori. È collegato alla vena caudale e passa in avanti attraverso il fegato al seno.
Negli embrioni di amnioti le vene addominali laterali sono note come vene ombelicali o allantoiche perché drenano l’allantois. Le vene vitelline e ombelicali si perdono alla nascita, solo i loro resti persistono.
Modificazioni delle vene:
Come nel caso delle arterie, le vene di diversi vertebrati sono disposte sullo stesso piano fondamentale. Tutte le variazioni mostrate da loro seguono una sequenza nella serie dei vertebrati. Nel suo sviluppo il sistema venoso dei vertebrati superiori passa attraverso gli stadi visti negli embrioni di forme inferiori.
1. Quando si forma il fegato, la parte prossimale delle vene vitelline o vene subintestinali forma le vene epatiche tra il fegato e il cuore, la parte distale della vena vitellina sinistra o vena subintestinale diventa la vena porta epatica che forma sinusoidi nel fegato per dare origine a un sistema portale epatico presente in tutti.
2. Le vene cardinali anteriori persistono come vene giugulari interne.
3. Tranne che nei pesci, le vene cardinali comuni diventano le vene precavali che entrano nel sinus venosus (anfibi, rettili) o nel padiglione auricolare destro (uccelli, mammiferi). Una vena succlavia si forma in ogni arto anteriore che si unisce alla vena precavale.
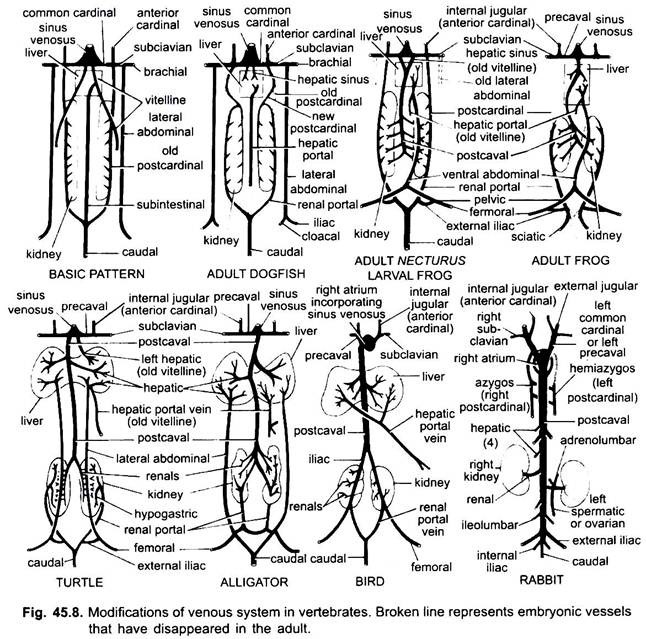
4. La vena caudale perde le sue connessioni con le vene subintestinali e subcardinali, la sua parte anteriore si divide in due rami che si uniscono alle vene cardinali posteriori in tutti tranne i mammiferi.
5. Vene cardinali posteriori persistono come tali pesci, ma in altri, ognuno si divide in due parti, una parte anteriore, che scompare in anfibi, rettili e uccelli, ma nei mammiferi il diritto porzione anteriore forme di una vena azygos, la sinistra può formare una hemiazygos vena che perde la sua connessione con il precaval e si unisce alla vena azygos da una trasversale anastomosi; il posteriore porzioni posteriori vene cardinali che hanno aderito alla vena caudale, diventare renale portale vene.
La vena porta renale di ciascun lato forma capillari nel rene per formare un sistema portale renale che è completo in pesci, anfibi e rettili, ma diventa molto ridotto negli uccelli ed è assente nei mammiferi perché le porzioni posteriori delle vene cardinali posteriori scompaiono nei mammiferi.
6. Le vene portale renali dei pesci raccolgono il sangue solo dalla coda, ma negli anfibi, nei rettili e negli uccelli usurpano anche le vene dalle gambe.
7. In alcuni pesci e in tutti i tetrapodi le vene vitelline si uniscono alla vena subcardinale per formare una vena postcavale. Negli anfibi e nei rettili la vena postcavale si estende alle estremità posteriori dei reni, negli uccelli si unisce alle vene portale renale, riducendo così il sistema portale renale, nei mammiferi la vena postcavale si unisce alle vene dalle gambe e dalla coda, in modo che il sistema portale renale sia completamente eliminato.
8. Le due vene addominali laterali persistono come tali nei pesci, ma da Dipnoi verso l’alto si fondono per formare una vena addominale anteriore che unisce la vena porta epatica vicino al fegato, la vena addominale anteriore, quindi, collega il portale renale e il sistema portale epatico. Una vena iliaca si forma in ogni arto posteriore che si unisce alla vena addominale anteriore. La vena addominale anteriore inizia a perdere la sua importanza nei rettili, negli uccelli viene modificata per formare le vene epigastriche e coccigeo-mesenteriche, nei mammiferi scompare, tranne che nel Tachyglossus.
9. Nelle forme di respirazione aerea, le vene polmonari crescono dal padiglione auricolare sinistro e entrano nei polmoni.
10. Le vene ombelicali (allantoiche) dell’embrione scompaiono quando i polmoni diventano funzionali come organi respiratori.
Tetrapoda:
Il sistema venoso degli anfibi assomiglia molto a quello dei lungfishes tranne che la vena addominale entra nel sistema portale epatico piuttosto che nel seno venoso. I cardinali anteriori persistono come vene giugulari interne in tutti i tetrapodi adulti. Le vene giugulari inferiori sono assenti. I cardinali comuni diventano la vena cavae anteriore o precavali, che si uniscono al seno venoso negli anfibi e nei rettili.
Negli uccelli e nei mammiferi, i precavali entrano direttamente nel padiglione auricolare destro del cuore, perché il seno venoso è assente. Nell’uomo, nel gatto, ecc., il precaval sinistro è perso così il sangue di lato sinistro entra nel precaval destro attraverso un ramo brachio-cephalic. Nelle salamandre senza polmoni, le vene polmonari sono assenti poiché il setto interatriale è incompleto.
Le vene post cardinali si uniscono alla vena caudale posteriormente e precavali anteriormente per formare i cardinali comuni nella rana larvale e nel Necturus.
Nei rettili, vi è un maggiore sviluppo delle vene polmonari e della vena postcavale e riduzione dell’importanza del sistema portale renale che porta il sangue ai reni dalla parte posteriore del corpo.
Negli uccelli, ci sono due vene precavali funzionali formate dall’unione della giugulare e della succlavia su ciascun lato e un post cavale completo. Riceve sangue dagli arti attraverso i portali renali, che passano attraverso i reni, ma non si rompono in capillari e non sono paragonabili ai portali renali dei vertebrati inferiori.
Nei mammiferi, ci possono essere una o due vene precavali. C’è una singola vena postcavale, il cui sviluppo embriologico è molto complicato. Tutte le vene cavali entrano direttamente nel padiglione auricolare destro, poiché il seno venoso viene assorbito nella parete di questa camera del cuore nella vita embrionale. Non esiste un sistema portale renale, sebbene il sistema portale epatico assomigli molto a quello di altri vertebrati.