PUBBLICITÀ:
In questo articolo discuteremo del sistema digestivo dei vertebrati con l’aiuto di diagrammi adatti.
Apparato digerente embrionale:
Archenteron:
L’archenteron embrionale diventa il rivestimento del tratto digestivo adulto e di tutti i suoi derivati. Il mesoderma splancnico aggiunge strati di tessuto connettivo e muscoli lisci intorno all’archenteron. L’invaginazione ectodermica della testa forma lo stomodeo che porta nella cavità orale e un’invaginazione ectodermica mid-ventrale simile forma il proctodeo, che porta nel hindgut.
PUBBLICITÀ:
Lo stomodeo diventa la cavità buccale adulta e dà origine allo smalto dei denti, alla copertura epiteliale della lingua, alle ghiandole, ad esempio mucose, veleni e salivari, ecc., e la sacca di Rathke di ghiandola pituitaria anteriore. Il proctodeo forma una piccola parte terminale della cloaca nei vertebrati inferiori e nel retto nei mammiferi.
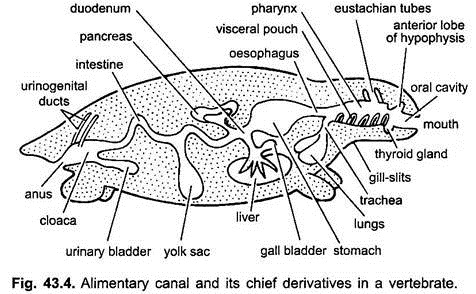
Il canale alimentare negli embrioni dallo stomaco alla cloaca è attaccato alla parete dorsale del corpo da una doppia piega del peritoneo, chiamata mesentere dorsale, e alla parete ventrale del corpo da un mesentere ventrale. Negli adulti, il mesentere dorsale persiste ma il mesentere ventrale scompare lasciando solo nella regione del fegato e della vescica urinaria.
Tratto digestivo dell’adulto:
Il tratto digestivo si differenzia per diverse funzioni nelle seguenti regioni: bocca, cavità buccale, faringe, esofago, stomaco, intestino tenue, intestino crasso e cloaca. Le seguenti escrescenze derivano dal tratto digestivo – ghiandole orali, sacca di Rathke, ghiandola tiroidea, fessure branchiali, cavità timpanica, timo e altre ghiandole di fessure branchiali, trachea, polmoni, vescica natatoria, fegato, pancreas, sacco vitellino e vescica urinaria.
PUBBLICITÀ:
Istologia:
La parete del canale alimentare è costituita da quattro strati concentrici.
Sono:
(i) Un peritoneo viscerale più esterno o un mantello sieroso è costituito da cellule mesoteliali e da un sottile strato di tessuto connettivo. Manca nell’esofago,
PUBBLICITÀ:
(ii) Al di sotto di questo è uno strato muscolare formato da fibre muscolari lisce disposte in fibre muscolari circolari longitudinali esterne e interne. Tra i due strati di muscoli è una rete di cellule nervose e fibre nervose del sistema nervoso autonomo, noto come plesso mienterico o plesso di Auerbach.
iii) Sotto lo strato muscolare si trova una sottomucosa costituita da tessuto connettivo con fibre elastiche, grasso, vasi sanguigni e linfatici, cellule nervose e ghiandole delle fibre,
iv) Lo strato più interno è una mucosa composta da tre regioni:
a) Mucosa muscularis esterna-più stretta delle fibre muscolari lisce longitudinali ed interne circolari esterne.
(b) Strato medio sottile di lamina propria di tessuto connettivo, vasi sanguigni, nervi e noduli di tessuto linfatico, e
(c) Una membrana basale che supporta uno strato di cellule epiteliali colonnari che sono spesso ghiandolari e ciliati.
Bocca:
Bocca è l’apertura che conduce nella cavità buccale. Nelle lamprede (ciclostomi)è un’apertura circolare alla base dell’imbuto buccale e rimane permanentemente aperta per mancanza di mascelle, ecc. Negli gnatostomi è terminale. La bocca è delimitata da labbra che sono immobili e formate da pelle cornificata in pesci, anfibi e rettili. Nei mammiferi questi sono carnosi e muscolosi.
Cavità buccale:
PUBBLICITÀ:
Lo spazio tra le labbra e le mascelle è un vestibolo. Può essere delimitato all’esterno dalle guance e all’interno dalle gengive. Le ghiandole mucose delle guance si aprono nel vestibolo. La bocca si apre in una cavità buccale, che è uno spazio tra la bocca e la faringe. Il punto esatto in cui l’ectoderma stomodeale e l’endoderma faringeo si fondono è variabile e non facile da discernere.
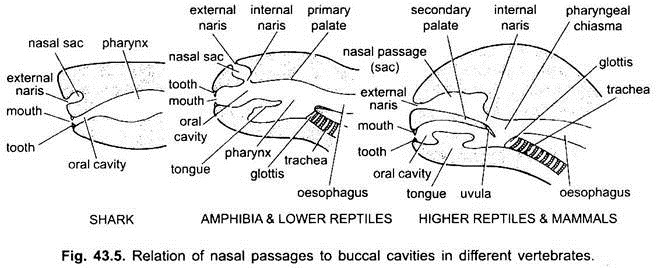
Negli elasmobranchi e nella maggior parte dei pesci ossei le cavità nasali non si aprono nella cavità buccale. In Chondrichthyes e tetrapoda (anfibi e la maggior parte dei rettili) la cavità nasale si aprono nella cavità buccale da choanae o interno delle narici, che sono primitivamente posizionato anteriormente, ma in coccodrilli, uccelli e mammiferi diventano posteriore della faringe dovuta alla formazione di un palato secondario, che di fatto separa l’apparato respiratorio nasale passaggio dalla cavità della bocca o il passaggio del cibo.
Negli uccelli, questo palato è una fessura a causa della quale le cavità nasali e buccali comunicano tra loro. Nei mammiferi, il palato secondario è continuato posteriormente come palato molle membranoso. Negli esseri umani il palato molle si blocca nella faringe laringea sotto forma di processo carnoso, chiamato uvula.
Derivati della cavità buccale:
1. Ghiandole orali:
Ci sono due tipi di ghiandole multicellulari tegumetarie che si aprono nella cavità buccale. Sono ghiandole mucose e ghiandole enzimatiche. I pesci e gli anfibi acquatici hanno solo ghiandole mucose. I rettili hanno ghiandole in gruppi, come le ghiandole palatine, linguali, sublinguali e labiali chiamate in base alla posizione, producono anche muco.
Nei serpenti velenosi le ghiandole labiali superiori sono modificate per secernere veleno, mentre nel mostro Gila le ghiandole sublinguali producono veleno. Gli uccelli hanno ghiandole sublinguali e una ghiandola nell’angolo della bocca. I mammiferi hanno molte piccole ghiandole mucose oltre alle ghiandole salivari vere e ingrandite che sono enzimatiche. Sono ghiandole salivari parotide, sublinguali, sottomascellari e infraorbitali, che secernono mucina e ptyalin.
2. Lingua:
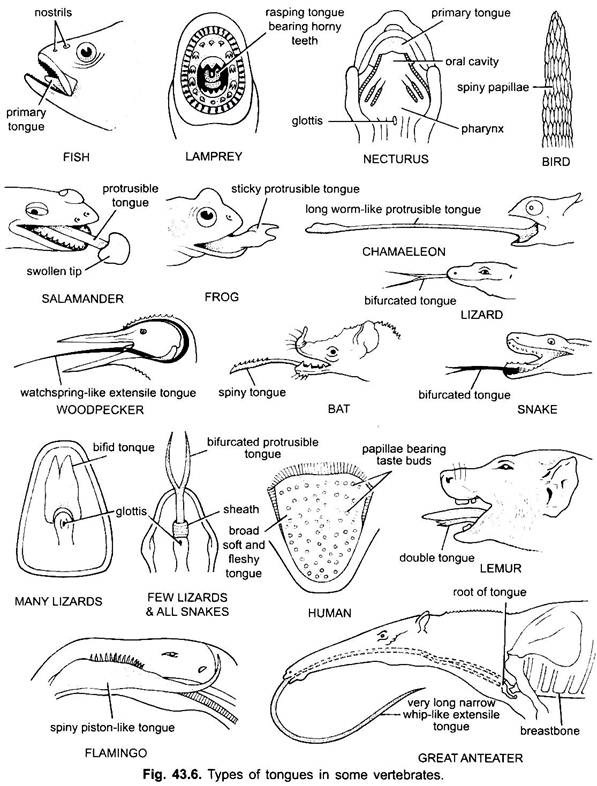
La lingua si trova principalmente in tutti i vertebrati. La lingua nei vertebrati mostra molta diversità e non è omologa. Nei ciclostomi, c’è una lingua muscolosa, carnosa e raspante con denti cornei per raspare la pelle e i muscoli della loro preda.
I pesci hanno una lingua primaria formata da una piega carnosa del pavimento buccale. Non ha muscoli, ma recettori e denti sono presenti sulla lingua in alcuni pesci ossei. La lingua è coperta da membrana mucosa. In alcuni anfibi la lingua è carente o immobile. La maggior parte degli anfibi, tuttavia, hanno una lingua protrusibile e in alcune rane e rospi può essere ripiegato su se stesso quando non in uso.
Può essere buttato fuori dalla bocca da un rapido afflusso di linfa per catturare gli insetti. La lingua nelle lucertole e nei serpenti è spesso molto sviluppata. Nei camaleonti, è molto estensibile usato per catturare insetti. La punta è ispessita e appiccicosa. La punta biforcuta della lingua nei serpenti serve come mezzo per trasferire stimoli chimici dall’ambiente esterno agli organi vomero-nasali accoppiati sul tetto della bocca.
Nelle tartarughe e nei coccodrilli, la lingua non può essere estesa. La lingua amniota ha muscoli volontari, riceve il nervo ipoglosso e ha ghiandole e papille gustative. Sviluppa anche muscoli intrinseci che muovono la lingua. Negli uccelli, la lingua è snella e ha una copertura cornea. In alcuni uccelli la lingua è immobile, mentre in alcuni uccelli è lunga, protrattile e spesso utilizzata per catturare il cibo.
Nella maggior parte dei mammiferi, ad eccezione delle balene, la lingua è altamente sviluppata e capace di notevoli movimenti, oltre all’estensione e alla retrazione, a causa della presenza di un numero di muscoli intrinseci.
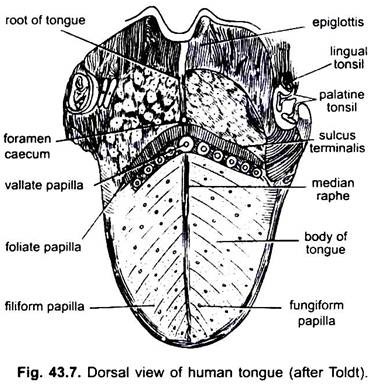
Nei mammiferi la mucosa sotto la lingua forma una piega mediana, chiamata frenulo che unisce la lingua al pavimento della bocca. Nei mammiferi la superficie superiore della lingua porta quattro tipi di papille, (filiforme, fungiforme, foliata e circumvallata), con papille gustative ad eccezione delle papille filiformi.
3. Denti:
I vertebrati hanno due tipi di denti attaccati alle ossa mascellari: denti epidermici e denti veri. I denti epidermici sono meglio sviluppati nei ciclostomi. Sono strutture dure, coniche e cornee derivate dallo strato corneo. Nelle lamprede si trovano sulle pareti dell’imbuto buccale e sulla lingua. Le larve di girino di rane e rospi hanno denti epidermici seghettati in file sulle labbra. Nei mammiferi l’ornitorinco adulto ha denti epidermici.
Denti veri:
I denti non si trovano nelle balene fanoni e nei formichieri nei mammiferi, e agnati, storioni, alcuni rospi, sirene, tartarughe e uccelli moderni, ecc. Nei vertebrati inferiori (come pesci, anfibi e la maggior parte dei rettili) i denti possono essere sostituiti continuamente un numero indefinito di volte, tali denti sono chiamati polifiodonti. Questi denti sono omodonti (tipo simile) e acrodonti. (con le ossa mascellari).
Nella maggior parte dei mammiferi i denti sono diphyodont, thecodont e heterodont. In alcuni mammiferi questi sono monofiodonti con un solo set di denti, ad esempio, talpe, scoiattolo indiano. I denti sono simili nella struttura alle squame placoidi degli squali, formate da una cavità pulpare centrale, attorno alla quale è presente uno strato spesso ma morbido, la dentina, che è ricoperta esternamente da uno smalto sottile ed estremamente duro. Si suppone che questi derivassero da squame ossee di ostracodermi e placodermi. Per i dettagli i lettori possono vedere la dentatura nei mammiferi.
4. Adenohypophysis:
Il lobo anteriore della ghiandola pituitaria si sviluppa come evaginazione dorsale dello stomodeo, chiamata sacca di Rathke, che si restringe per formare i lobi anteriori e medi della ghiandola pituitaria (adenohypophysis). Il lobo posteriore dell’ipofisi o neuroipofisi è l’evaginazione ventrale del diencefalo, chiamato infundibolo. Quindi, è parte nervosa.
Faringe:
La parte del canale alimentare immediatamente dietro la cavità buccale è una faringe, rivestita di endoderma. È un passaggio comune che serve sia per la digestione che per la respirazione. Come parte del sistema digestivo è usato come passaggio per il cibo dalla cavità buccale all’esofago, i suoi muscoli iniziano la deglutizione.
Nei pesci la faringe è grande e perforata lateralmente per fessure branchiali, mentre nei tetrapoda è corta e porta aperture delle narici. Negli embrioni, la parete della faringe emana un numero di evaginazioni che si sviluppano in spiracoli, fessure branchiali, vesciche d’aria, polmoni, tonsille e alcune ghiandole endocrine (ad esempio, timo, tiroide e paratiroidi).
Esofago:
L’esofago è corto nella maggior parte dei pesci e degli anfibi perché privo di collo, ma è più lungo negli amnioti per presenza di collo. L’esofago dei rettili è più allungato di quello dei pesci e degli anfibi. Negli uccelli granivori e carnivori, una parte dell’esofago viene ingrandita in una sacca simile a un sacco chiamata coltura che serve a conservare il cibo che è stato mangiato rapidamente.
Il raccolto è essenzialmente privo di ghiandole digestive anche se nei piccioni il raccolto ha 2 ghiandole del raccolto in entrambi i sessi, in realtà non sono ghiandole ma strutture che formano cellule, le cellule formano “latte di piccione” che viene alimentato ai giovani. Nei mammiferi, l’esofago è lungo, privo di ghiandole e varia in relazione alla lunghezza del collo.
Passa attraverso il diaframma, la porzione sotto il diaframma è ricoperta da peritoneo viscerale che manca dalla parte superiore. L’esofago ha ghiandole mucose. Il suo rivestimento forma pieghe longitudinali, o papille carnose simili a dita (elasmobranchi) o papille cornee nelle tartarughe marine.
Istologicamente, l’esofago differisce dal resto del canale alimentare in tre fatti:
(i) Non ha peritoneo viscerale perché si trova al di fuori del celoma, il suo strato di copertura più esterno è una sottile tunica avventizia.
(ii) Le fibre muscolari nella sua parte anteriore sono striate, la parte centrale ha muscoli sia striati che non striati, e la parte posteriore ha solo muscoli non striati. Ma ci sono eccezioni nel ruminare i mammiferi, tutti i muscoli sono a strisce o volontari.
(iii) Il rivestimento della membrana mucosa è costituito da cellule epiteliali squamose stratificate e non da cellule colonnari.
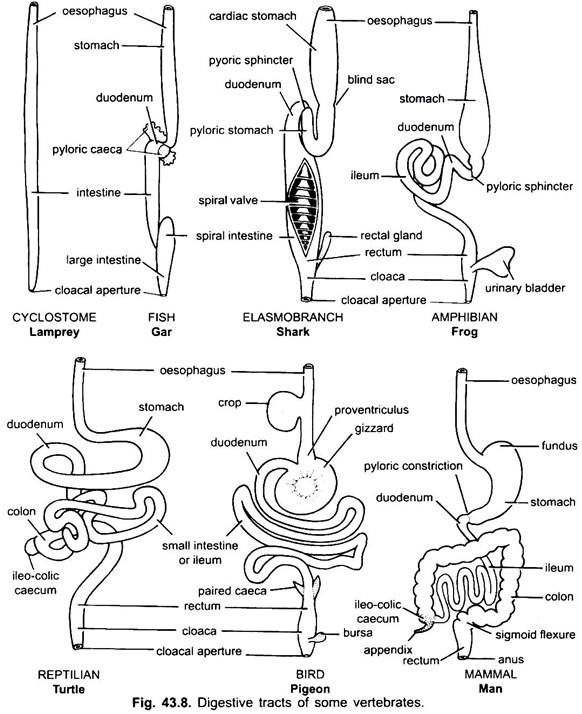
Stomaco:
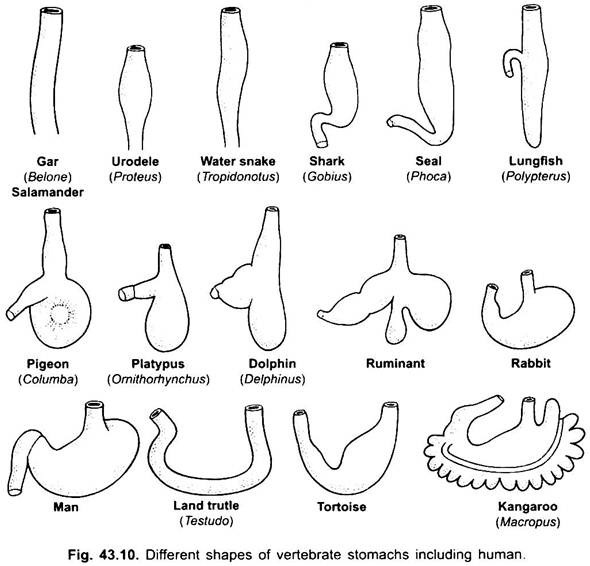
Non c’è praticamente stomaco nei ciclostomi, chimere, pesci polmonari e alcuni pesci teleostei primitivi, poiché non ha ghiandole gastriche, ma nella maggior parte dei pesci e dei tetrapodi è dilatato per la conservazione e la macerazione del cibo solido e la digestione del cibo perché contiene ghiandole gastriche.
La prima parte dello stomaco, accanto all’esofago, è la regione cardiaca e l’estremità inferiore vicino all’intestino è la regione pilorica, che ha un piloro o una valvola pilorica in cui il rivestimento della mucosa è circondato da uno spesso muscolo sfintere che regola l’apertura e la chiusura dello stomaco pilorico nell’intestino.
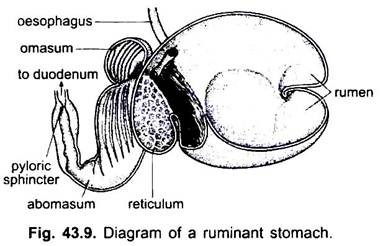
Lo stomaco è dritto nei ciclostomi, gar, Belone,ecc., e a forma di fuso in Proteus, Necturus, alcune lucertole e serpenti. Nelle tartarughe e nelle tartarughe, è un ampio tubo curvo, e negli elasmobranchi lo stomaco è a forma di J. Nei coccodrilli e negli uccelli lo stomaco ha due parti, un proventricolo con ghiandole gastriche e un ventriglio altamente muscolare, che rappresenta la regione pilorica e ha un rivestimento duro e cornificato per macinare il cibo.
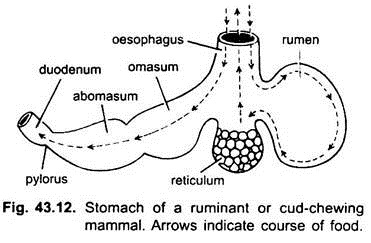
Nei mammiferi lo stomaco giace trasversalmente e può essere un semplice sacco o diviso in 3 regioni, vale a dire cardiaca, fundica e pilorica e ogni regione ha le sue ghiandole gastriche. In molti ruminanti lo stomaco ha quattro camere: un rumine, un reticolo, un omaso e un abomaso. Si sostiene che le prime tre camere sono modifiche dell’esofago, e abomaso è il vero stomaco che rappresenta il cuore, fundica, e parti piloriche dello stomaco.
È stato dimostrato embriologicamente che tutte e quattro le camere sono regioni modificate dello stomaco. Nei cammelli, non c’è omaso, il rumine e il reticolo hanno cellule d’acqua simili a sacche che una volta si credeva immagazzinassero acqua, ma probabilmente sono digestive.
Istologicamente lo stomaco presenta le tipiche parti del canale alimentare, ma presenta due peculiarità, la mucosa muscularis è costituita da uno strato longitudinale esterno e uno strato circolare interno di muscoli. Il rivestimento dell’epitelio è spesso con diversi tipi di cellule ghiandolari che formano ghiandole gastriche di tre tipi chiamati ghiandole gastriche cardiache, fundiche e piloriche.
Le ghiandole cardiache e piloriche secernono solo muco dalle loro cellule superficiali. Le ghiandole fundiche (o ghiandole cardiache in alcuni) hanno tre tipi di cellule, le cellule del collo mucoso producono muco, le cellule ossintiche producono acido cloridrico, possono essere presenti anche nella regione cardiaca, le cellule zimogene o le cellule peptiche secernono pepsina.
Nella maggior parte degli animali le cellule di zimogeno secernono anche due proenzimi chiamati propepsina e prorennina che vengono convertiti dall’acido cloridrico rispettivamente in pepsina e rennina. Le secrezioni di tutte le cellule dello stomaco formano una miscela chiamata succo gastrico.
Intestino tenue:
L’intestino tenue è un tubo lungo, stretto e arrotolato dopo il piloro. È la parte più importante del tratto digestivo perché la digestione e l’assorbimento del cibo avvengono in esso. Nei ciclostomi, l’intestino è un breve tubo dritto con un lembo longitudinale disposto a spirale che si estende in esso.
In elasmobranchs, è diviso in piccole e grandi porzioni e la piccola porzione ha una valvola a spirale che aumenta notevolmente la superficie assorbente. Una valvola a spirale è presente anche nell’intestino tenue di alcuni pesci ossei più primitivi, ma manca di forme superiori in cui l’intestino è lungo e arrotolato.
Nelle cecilie, è poco arrotolato e non differenziato in un tratto piccolo e grande. Nelle rane e nei rospi è relativamente lungo e arrotolato. Nei rettili, è più arrotolato che negli anfibi. Per la prima volta nei vertebrati si verifica un cieco o un diverticolo cieco all’incrocio tra intestino tenue e crasso.
Tuttavia, questo non è permanente in tutti i rettili. Negli uccelli, l’intestino tenue è arrotolato o avvolto e una o due coliche caeca sono presenti anche all’incrocio tra intestino tenue e crasso. Nella maggior parte dei mammiferi anche l’intestino tenue è proporzionalmente lungo e arrotolato. La sua lunghezza, tuttavia, è correlata con le abitudini alimentari. Negli erbivori è relativamente più lungo rispetto agli insettivori e ai carnivori.
C’è una tasca cieca o cieco alla giunzione del colon e dell’intestino tenue che è generalmente piccola nelle specie carnivore e piuttosto lunga in molti erbivori. La prima parte dell’intestino tenue è il duodeno, che è breve a partire dal piloro e termina oltre l’ingresso dei dotti pancreatici ed epatici.
Ha molti villi piegati e contiene le ghiandole di Brunner ramificate nella sottomucosa che secernono muco, alcuni liquidi acquosi alcalini e un piccolo enzima. Il duodeno produce anche due ormoni chiamati secretina e colecistochinina che stimolano il pancreas e la cistifellea a liberare i loro succhi. I dotti della cistifellea e del pancreas si aprono nel duodeno.
Dietro il duodeno c’è un ileo, che solo nei mammiferi è differenziato in un digiuno più piccolo anteriore e un ileo più lungo posteriore. Un gran numero di piccole ghiandole digestive sono presenti nell’intestino tenue. Sono ghiandole tubulari o cripte di Lieberkuhn trovati attraverso l’intera lunghezza, secernono muco e un succus entericus che ha diversi enzimi.
Il rivestimento dell’intestino tenue viene piegato per formare piccoli villi, che aumentano l’area superficiale per la secrezione e l’assorbimento. I villi sono coperti densamente da minuscole proiezioni simili a dita, chiamate microvilli che aiutano nell’assorbimento nei villi. Nei mammiferi si trovano noduli di tessuto linfoide chiamati cerotti di Peyer sull’ileo.
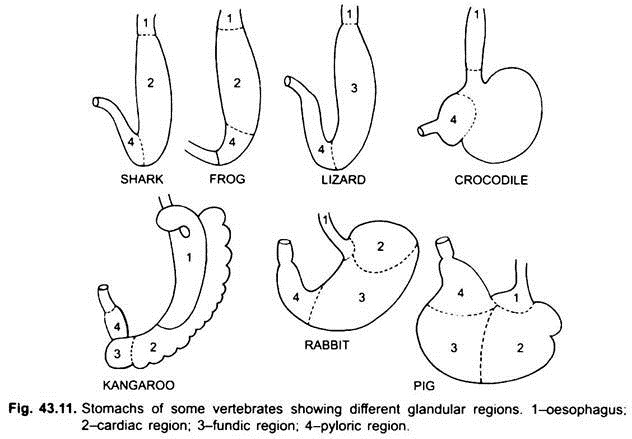
Intestino crasso:
L’intestino crasso ha un diametro maggiore rispetto all’intestino tenue. È generalmente corto in pesci, anfibi, rettili e uccelli, ma nei mammiferi è lungo. Nelle forme inferiori l’intestino crasso forma un retto, ma in tetrapoda ha un colon e un retto terminale. Nella maggior parte dei pesci e degli anfibi, la parte terminale del retto conduce in una cloaca formata dal proctodeo.
Il retto, i dotti escretori e i dotti genitali si aprono nella cloaca e si apre all’esterno da un’apertura cloacale. Ma in molti pesci ossei e in tutti i mammiferi (eccetto i monotremi) il retto e i dotti urinogenitali hanno aperture separate verso l’esterno; l’apertura del primo è un ano.
Il retto dei mammiferi non è omologo con il retto dei vertebrati poiché nei mammiferi è derivato dal partizionamento della cloaca embrionale. Nella maggior parte degli embrioni vertebrati c’è un intestino postanale come estensione dell’intestino nella coda, ma scompare in seguito.
In elasmobranchs l’intestino crasso porta un paio di ghiandole rettali che secernono muco e cloruro di sodio. Negli amnioti c’è una valvola ileocolica alla giunzione di intestino tenue e crasso, che è assente nei pesci. Impedisce ai batteri di entrare nell’ileo dal colon.
Negli amnioti da questa giunzione nasce un cieco ileocolico che è due negli uccelli. Contiene batteri che digeriscono la cellulosa. È molto lungo nei mammiferi erbivori (coniglio, cavallo, mucca, ecc.). Nei primati il cieco è piccolo con un’appendice vermiforme vestigiale.
Ghiandole digestive:
1. Fegato:
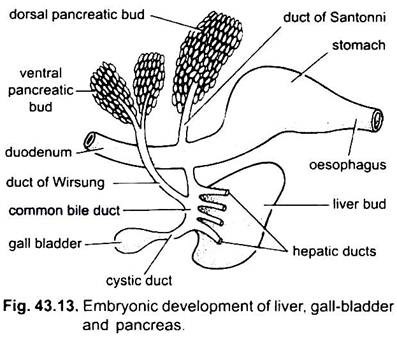
Il fegato si presenta come una singola o doppia escrescenza dalla parete ventrale dell’archenteron embrionale. Questa escrescenza forma un diverticolo epatico cavo, che presto si differenzia in una parte anteriore, che prolifera per diventare il fegato e i suoi dotti biliari, e una parte posteriore, che dà origine alla cistifellea e al dotto cistico. I dotti biliari si uniscono per formare un dotto epatico che si unisce con il dotto cistico per formare un dotto biliare comune o dotto choledochus. La regione dell’archenteron da cui nasce il fegato diventa il duodeno.
Il fegato è la più grande ghiandola lobata del corpo, sospesa da un doppio strato di peritoneo dal setto trasversale o dal suo rappresentante.
Una cistifellea è per la conservazione della bile secreta dalle cellule epatiche, si trova nel fegato e drena nel duodeno attraverso il dotto biliare comune formato dall’unione del dotto cistico e del dotto epatico. Una cistifellea non è indispensabile e manca in molti uccelli e mammiferi.
Un fegato è presente in tutti i vertebrati. Nei ciclostomi, è piccolo, singolo lobato (lamprede) e due lobati in hagfishes. È bilobato in elasmobranchi, due o tre lobati in pesci ossei, anfibi, rettili e uccelli e molti lobati nei mammiferi. Il fegato è lungo, stretto e cilindrico nei pesci, negli urodeli e nei serpenti.
È corto, largo e appiattito negli uccelli e nei mammiferi. Una cistifellea e dotto biliare sono presenti nei ciclostomi larvali, ma sono assenti nell’adulto. Pesci, anfibi e rettili hanno generalmente una cistifellea, ma manca in molti uccelli. La maggior parte dei mammiferi ha una cistifellea, ma è assente nei Cetacei e negli Ungulati.
Il fegato secerne una bile acquosa e alcalina ma non ha enzimi. Neutralizza l’acidità del cibo che entra nel duodeno. Aiuta nella digestione dei grassi.
2. Pancreas:
Il pancreas è formato dall’endoderma dell’archenteron embrionale. Un singolo diverticolo dorsale dal duodeno embrionale e una o due escrescenze ventrali dal fegato formano diverticoli pancreatici. Le parti prossimali dei diverticoli formano dotti pancreatici, ma questi dotti subiscono una riduzione o fusione in modo che solo uno o due dotti pancreatici rimangano nell’adulto, si aprono nel duodeno separatamente o dopo l’unione con il dotto biliare comune.
Le parti distali dei diverticoli subiscono il germogliamento per formare la massa principale delle cellule pancreatiche a cui vengono aggiunti i derivati mesodermici. Quindi, viene creata una singola ghiandola che ha diversi lobi che formano un pancreas diffuso o compatto.
Il pancreas è sia una ghiandola esocrina che endocrina, legata insieme da delicati fili di tessuto connettivo. La parte esocrina secerne enzimi digestivi che vengono versati nel duodeno attraverso dotti pancreatici. Mentre la parte endocrina secerne ormoni come l’insulina e il glucagone.
Un pancreas è presente in tutti i vertebrati. Nelle lamprede, in alcuni pesci ossei, nei lungfish e nei tetrapodi inferiori, è un organo diffuso incorporato nel fegato, nei mesenteri e nella parete intestinale. Hagfishes hanno un piccolo pancreas. Gli elasmobranchi hanno un pancreas bilobato ben definito. In tetrapoda superiore è generalmente una ghiandola compatta. Uno o due dotti pancreatici si aprono nel duodeno.