annonser:
i denna artikel kommer vi att diskutera om matsmältningssystemet hos ryggradsdjur med hjälp av lämpliga diagram.
embryonal matsmältningskanal:
Archenteron:
den embryonala archenteronen blir slemhinnan i den vuxna matsmältningskanalen och av alla dess derivat. Splanchnic mesoderm lägger till lager av bindväv och släta muskler runt archenteron. Ektodermal invagination av huvudet bildar stomodaeum som leder in i munhålan, och en liknande Mid-ventral ektodermal invagination bildar proctodaeum, vilket leder in i bakgut.
annonser:
stomodaeum blir den vuxna bukkala håligheten och ger upphov till tandemalj, epitelbeläggning av tunga, körtlar, t.ex. slemhinnor, gift och saliv, etc. och Rathkes påse med främre hypofysen. Proctodaeum bildar antingen en liten terminal del av cloaca i nedre ryggradsdjur och rektum hos däggdjur.
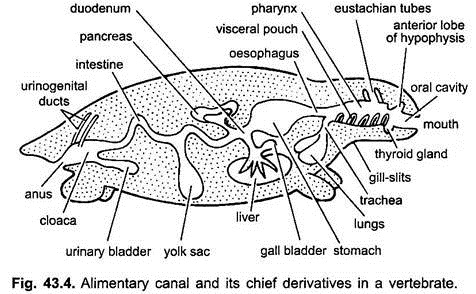
matsmältningskanalen i embryon från Mage till cloaca är fäst vid ryggkroppsväggen genom en dubbelvikt av bukhinnan, kallad dorsal mesenteri, och till ventral kroppsvägg med en ventral mesenteri. Hos vuxna kvarstår dorsal mesenteri men ventral mesenteri försvinner och lämnar endast i regionen lever och urinblåsa.
mag-tarmkanalen hos vuxna:
matsmältningskanalen skiljer sig åt för olika funktioner i följande regioner – mun, buckalhålighet, svalg, matstrupe, mage, tunntarmen, tjocktarmen och cloaca. Följande utväxter uppstår från mag-tarmkanalen – munkörtlar, rathkes påse, sköldkörtel, gillklyftor, tympanisk hålighet, tymus och andra körtlar av gillklyftor, luftstrupen, lungor, simblåsa, lever, bukspottkörtel, äggula och urinblåsa.
annonser:
histologi:
väggen i matsmältningskanalen är gjord av fyra koncentriska lager.
de är:
(i) en yttersta visceral peritoneum eller serös päls är gjord av mesotelceller och tunt skikt av bindväv. Det saknas i matstrupen,
annonser:
(ii) under detta finns ett muskulärt skikt bildat av glatta muskelfibrer anordnade i yttre längsgående och inre cirkulära muskelfibrer. Mellan de två muskelskikten finns ett nätverk av nervceller och nervfibrer i det autonoma nervsystemet, känt som myenterisk plexus eller plexus av Auerbach.
(iii) under muskelskiktet finns en submukosa gjord av bindväv med elastiska fibrer, fett, blod och lymfkärl, nervceller och fibrer körtlar,
(iv) det innersta skiktet är en slemhinna som består av tre regioner:
(a) yttre smal muscularis slemhinna av yttre längsgående och inre cirkulära glatta muskelfibrer.
(b) mitt tunt lager av lamina propria av bindväv, blodkärl, nerver och knölar av lymfatisk vävnad, och
(c) ett källarmembran som stöder ett lager av kolumna epitelceller som ofta är körtel och cilierade.
mun:
mun är öppningen som leder in i buckalhålan. I lampreys (cyklostomer) är det en cirkulär öppning vid basen av buckaltratten och förblir permanent öppen på grund av brist på käkar etc. I gnathostomer är det terminal. Munnen är avgränsad av läppar som är orörliga och bildade av kornad hud i fiskar, amfibier och reptiler. Hos däggdjur är dessa köttiga och muskulösa.
Buckalhålan:
annonser:
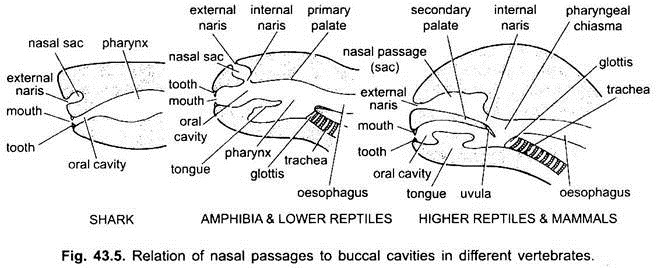
utrymmet mellan läpparna och käftarna är en vestibul. Det kan begränsas på utsidan av kinderna och på insidan av tandköttet. Slemhinnor i kinderna öppnar sig i förbenet. Munnen öppnar sig i en buckalhålighet, som är ett utrymme mellan munnen och svalget. Den exakta punkten där stomodaeal ectoderm och pharyngeal endoderm sammanfogar är variabel och inte lätt att urskilja.
i elasmobranchs och de flesta beniga fiskar öppnar näshålorna inte in i buckalhålan. I Chondrichthyes och tetrapoda (amfibier och de flesta reptiler) öppnar näshålorna i buckalhålan av choanae eller inre nares, som är primitivt placerade främre, men i krokodiler, fåglar och däggdjur blir de bakre i struphuvudet på grund av bildandet av en sekundär gom, som effektivt separerar andningsnäspassagen från munhålan eller matpassagen.
hos fåglar är denna gom kluven på grund av vilka nasala och buckala hålrum kommunicerar med varandra. Hos däggdjur fortsätter sekundär gom bakåt som en membranös mjuk gom. Hos människor hänger mjuk gom i struphuvudet i form av köttig process, kallad uvula.
derivat av Buckalhålan:
1. Orala körtlar:
det finns två typer av integumetära flercelliga körtlar som öppnar sig i buckalhålan. De är slemhinnor och enzymatiska körtlar. Fiskar och vattenlevande amfibier har bara slemhinnor. Reptiler har körtlar i grupper, såsom palatin, lingual, sublingual och labial körtlar namngivna efter plats, de producerar också slem.
i giftiga ormar modifieras de övre labialkörtlarna för att utsöndra gift, medan de sublinguala körtlarna i Gila-monsteret producerar gift. Fåglar har sublinguala körtlar och en körtel i munvinkeln. Däggdjur har många små slemhinnor förutom sanna och förstorade spottkörtlar som är enzymatiska. De är parotid, sublingual, submaxillär och infraorbital spottkörtlar, utsöndrar mucin och ptyalin.
2. Tunga:
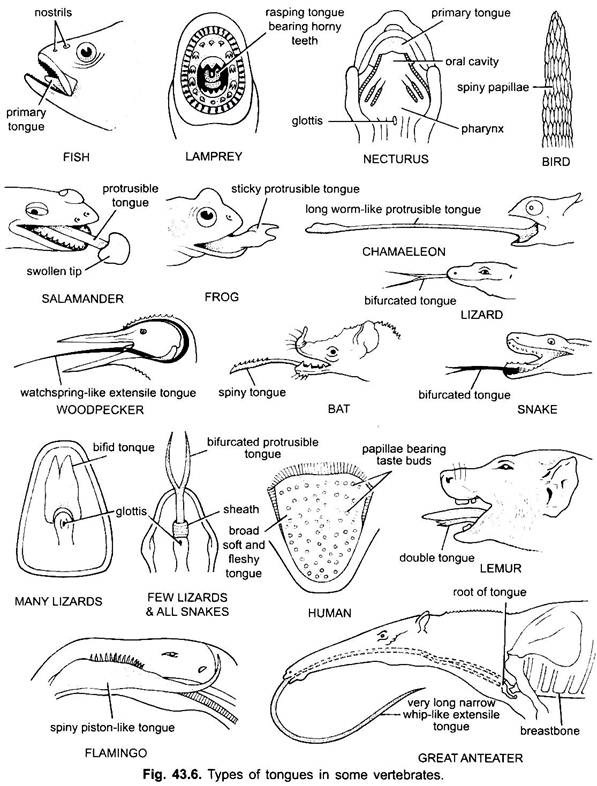
tungan finns mestadels i alla ryggradsdjur. Tungan hos ryggradsdjur visar mycket mångfald och är inte homologa. I cyklostomer finns en muskulös, köttig, raspande tunga med kåta tänder för att raspa huden och musklerna i sitt byte.
fiskar har en primär tunga som bildas av en köttig vik av buccal golvet. Det har inga muskler, men receptorer och tänder finns på tungan i vissa beniga fiskar. Tungan är täckt med slemhinnor. I vissa amfibier är tungan antingen saknad eller obeveklig. De flesta amfibier har dock en utskjutande tunga och i vissa grodor och paddor kan den vikas tillbaka på sig själv när den inte används.
det kan kastas ut ur munnen genom snabb inflöde av lymf för att fånga insekterna. Tungan i ödlor och ormar är ofta högt utvecklad. I kameleoner är det mycket utdragbart för att fånga insekter. Spetsen är förtjockad och klibbig. Den gafflade spetsen av tungan i ormar tjänar som ett sätt att överföra kemiska stimuli från den yttre miljön till de parade vomero-nasala organen på munnen.
i sköldpaddor och krokodiler kan tungan inte förlängas. Amniote-tungan har frivilliga muskler, den tar emot hypoglossalnerven och har körtlar och smaklökar. Det utvecklar också inneboende muskler som rör tungan. Hos fåglar är tungan smal och har en kåt täckning. I vissa fåglar är tungan orörlig, medan den i vissa fåglar är lång, långdragen och ofta används för att fånga maten.
i de flesta däggdjur, utom valar, är tungan högt utvecklad och kapabel till betydande rörelse, förutom förlängning och retraktion, på grund av närvaron av ett antal inneboende muskler.
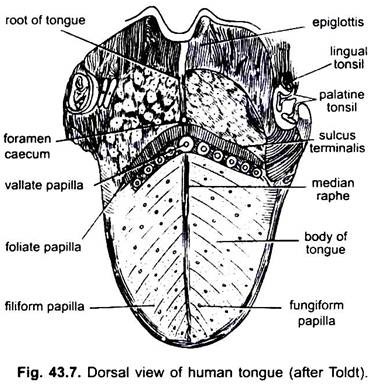
hos däggdjur bildar slemhinnan under tungan en medianveck, kallad frenulum som förenar tungan till munnen. Hos däggdjur har den övre ytan av tungan fyra typer av papiller, (filiform, fungiform, foliate och circumvallate), med smaklökar utom filiform papiller.
3. Tänder:
ryggradsdjur har två typer av tänder fästa vid käftben – epidermala tänder och sanna tänder. Epidermala tänder utvecklas bäst i cyklostomer. De är hårda, koniska, kåta strukturer som härrör från stratum corneum. I lampreys finns de på väggarna i buckaltratten och på tungan. Tadpole larver av grodor och paddor har tandade epidermala tänder i rader på läpparna. Hos däggdjur har den vuxna duckbill platypus epidermala tänder.
sanna tänder:
tänder finns inte i baleenvalar och anteaters hos däggdjur, och agnathans, störar, några paddor, sirener, sköldpaddor och moderna fåglar etc. I lägre ryggradsdjur (såsom fisk, amfibier och de flesta reptiler) tänder kan ersättas kontinuerligt ett obestämt antal gånger, sådana tänder kallas polyphyodont. Dessa tänder är homodont (liknande typ) och akrodont. (med käftbenen).
i de flesta däggdjur är tänderna diphyodont, thecodont och heterodont. I vissa däggdjur är dessa monophyodont som bara har en uppsättning tänder, t.ex. mol, Indisk ekorre. Tänderna är liknande i struktur till hajarnas placoidskalor, bildade av en central massahålighet, runt vilken det finns ett tjockt men mjukt skikt, dentinen, som är externt täckt av en tunn, extremt hård emalj. Dessa ska ha härletts från beniga skalor av ostracoderms och placoderms. För detaljer kan läsare se tandprotesen hos däggdjur.
4. Adenohypophysis:
den främre hypofysen utvecklas som en dorsal evagination av stomodaeum, kallad Rathkes påse, som constricts off för att bilda de främre och mellersta loberna i hypofysen (adenohypophysis). Den bakre loben av hypofysen eller neurohypophysis är den ventrala evaginationen av diencephalon, kallad infundibulum. Således är det nervös del.
svalg:
den del av matsmältningskanalen som ligger strax bakom buckalhålan är en svalg, fodrad med endoderm. Det är en vanlig passage som tjänar både för matsmältning och andning. Som en del av matsmältningssystemet används den som en passage för mat från buckalhålan till matstrupen, dess muskler initierar sväljning.
hos fiskar är svalget stort och perforerat i sidled för gillslitsar, medan det i tetrapoda är kort och bär näsborrar. I embryon avger svalgväggen ett antal evaginationer som utvecklas till spiracles, gill-clefts, luftblåsor, lungor, tonsiller och några endokrina körtlar (t.ex. tymus, sköldkörtel och parathyroider).
matstrupe:
matstrupen är kort i de flesta fiskar och amfibier eftersom de saknar nacke, men det är längre i amnioter på grund av närvaro av nacke. Matstrupen hos reptiler är mer långsträckt än hos fiskar och amfibier. Hos granivorösa och köttätande fåglar förstoras en del av matstrupen till en säckliknande påse som kallas gröda som tjänar till att lagra mat som har ätits snabbt.
grödan saknas i huvudsak i matsmältningskörtlar även om grödan i duvor har 2 grödor i båda könen, de är verkligen inte körtlar utan cellbildande strukturer, cellerna bildar ’duvmjölk’ som matas till unga. Hos däggdjur är matstrupen lång, saknar körtlar och varierar i förhållande till halsens längd.
den passerar genom membranet, delen under membranet är täckt med visceral peritoneum som saknas från övre delen. Esofagus har slemhinnor. Dess foder bildar längsgående veck, eller fingerliknande köttiga papiller (elasmobranchs) eller kåta papiller i havssköldpaddor.
histologiskt skiljer sig matstrupen från resten av matsmältningskanalen i tre fakta:
(i) den har ingen visceral bukhinnan eftersom den ligger utanför coelom, dess yttersta täckskikt är en tunn tunica adventitia.
(ii) muskelfibrerna i dess främre del är randiga, mellersta delen har både randiga och ostripade muskler och den bakre delen har endast ostripade muskler. Men det finns undantag i idisslande däggdjur, alla muskler är randiga eller frivilliga.
(iii) slemhinnan är gjord av stratifierade skivepitelceller och inte av kolonnceller.
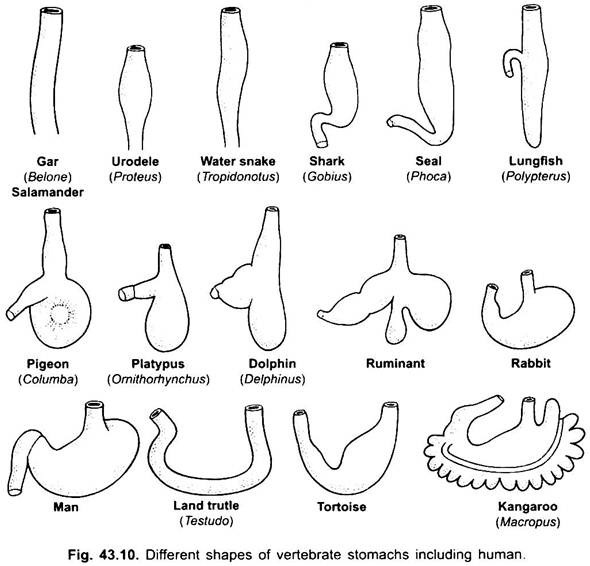
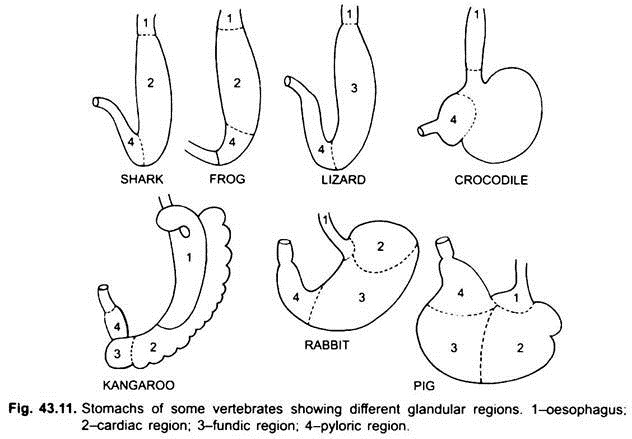
Mage:
det finns praktiskt taget ingen mage i cyklostomer, chimärer, lungfiskar och några primitiva teleostfiskar, eftersom den inte har några magkörtlar, men i de flesta fiskar och tetrapoda utvidgas den för lagring och maceration av fast mat och matsmältning eftersom den innehåller magkörtlar.
den första delen av magen, bredvid matstrupen, är hjärtregionen och den nedre änden nära tarmen är den pyloriska regionen, som har en pylorus eller pylorisk ventil i vilken slemhinnan är omgiven av en tjock sfinktermuskel som reglerar öppningen och stängningen av pylorisk mage i tarmen.
magen är rak i cyklostomer, gar, Belone, etc., och spindelformad i Proteus, Necturus, några ödlor och ormar. I sköldpaddor och sköldpaddor är det ett brett krökt rör, och i elasmobranchs är magen J-formad. I krokodiler och fåglar har magen två delar, en proventriculus med magkörtlar och en mycket muskulös Krås, som representerar den pyloriska regionen och har ett hårt, kornat foder för slipning av mat.
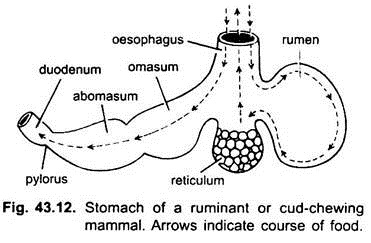
hos däggdjur ligger magen tvärs och kan vara en enkel säck eller uppdelad i 3 regioner, nämligen hjärt -, fundisk och pylorisk och varje region har sina magkörtlar. I många idisslare har magen fyra kamrar-en vom, retikulum, omasum och ett abomasum. Det hävdas att de tre första kamrarna är modifieringar av matstrupen, och abomasum är den sanna magen som representerar hjärt -, fundic-och pyloriska delar av magen.
det har visats embryologiskt att alla fyra kamrarna är modifierade regioner i magen. I kameler finns det ingen omasum, vommen och retikulum har påsliknande vattenceller som en gång tros lagra vatten, men de är förmodligen matsmältnings.
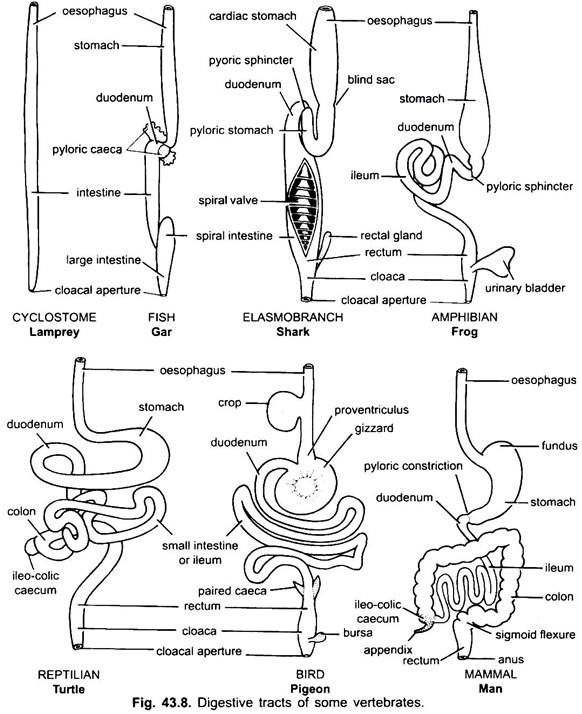
histologiskt har magen de typiska delarna av matsmältningskanalen, men den har två särdrag, muscularis slemhinna är gjord av ett yttre längsgående lager och ett inre cirkulärt lager av muskler. Epitelfodret är tjockt med flera typer av körtelceller som bildar magkörtlar av tre typer som kallas hjärt -, fundic-och pyloriska magkörtlar.
hjärt-och pylorkörtlarna utsöndrar endast slem från sina ytceller. Fundic körtlar (eller hjärtkörtlar i vissa) har tre typer av celler, slemhinnor halsceller producerar slem, oxyntiska celler producerar saltsyra, de kan vara närvarande i hjärtat regionen också, zymogen celler eller peptiska celler utsöndrar pepsin.
hos de flesta djur utsöndrar zymogencellerna också två proenzymer som kallas propepsin och prorennin som omvandlas av saltsyra till pepsin respektive rennin. Sekretionerna i alla magceller bildar en blandning som kallas magsaft.
Tunntarmen:
tunntarmen är lång, smal och lindad rör efter pylorus. Det är den viktigaste delen av matsmältningskanalen eftersom matsmältningen och absorptionen av mat äger rum i den. I cyklostomer är tarmen ett kort rakt rör med en spiralformad längsgående klaff som sträcker sig in i den.
i elasmobranchs är den uppdelad i små och stora delar, och den lilla delen har en spiralventil som kraftigt ökar absorberande ytan. En spiralventil finns också i tunntarmen hos några mer primitiva Benfiskar, men saknas i högre former där tarmen är lång och lindad.
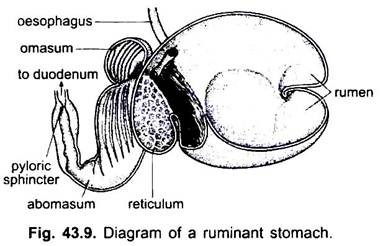
i caecilians är det lite lindat och inte differentierat till ett litet och stort område. Hos Grodor och paddor är den relativt lång och lindad. I reptiler är det mer spolat än i amfibier. För första gången hos ryggradsdjur uppstår en cecum eller blind divertikulum vid korsningen av små och stora tarmar.
detta är dock inte permanent i alla reptiler. Hos fåglar lindas tunntarmen eller slingras och en eller två kolikcaeca finns också vid korsningen mellan tunn-och tjocktarmen. I de flesta däggdjur är också tunntarmen proportionellt lång och lindad. Dess längd är dock korrelerad med matvanor. Hos växtätare är det relativt längre jämfört med insektsätare och köttätare.
det finns en blind ficka eller blindtarmen vid korsningen av tjocktarmen och tunntarmen som i allmänhet är liten i köttätande arter och ganska lång i många växtätare. Den första delen av tunntarmen är tolvfingertarmen, som är kort början från pylorus och slutar bortom ingången till bukspottkörtel-och leverkanaler.
den har många vikta villi och innehåller förgrenande Brunners körtlar i submukosa som utsöndrar slem, lite alkalisk vattnig vätska och ett litet enzym. Duodenum producerar också två hormoner som kallas secretin och cholecystokinin som stimulerar bukspottkörteln och gallblåsan att befria sina juicer. Kanaler från gallblåsan och bukspottkörteln öppnas i tolvfingertarmen.
bakom duodenum är en ileum, som endast hos däggdjur differentieras till en främre mindre jejunum och bakre längre ileum. Ett stort antal små matsmältningskörtlar finns i tunntarmen. De är rörformiga körtlar eller krypter av Lieberkuhn som finns genom hela längden, de utsöndrar slem och en succus entericus som har flera enzymer.
tunntarmens foder viks för att bilda små villi, vilket ökar ytan för utsöndring och absorption. Villi täcks tätt av små fingerliknande utsprång, kallade mikrovilli som hjälper till att absorberas i villi. Hos däggdjur finns knölar av lymfoid vävnad som kallas Peyers fläckar på ileum.
tjocktarmen:
tjocktarmen har en större diameter än tunntarmen. Det är i allmänhet kort i fiskar, amfibier, reptiler och fåglar, men hos däggdjur är det långt. I lägre former bildar tjocktarmen en rektum, men i tetrapoda har den en kolon och terminal rektum. I de flesta fiskar och amfibier leder den slutliga delen av ändtarmen till en cloaca som bildas av proctodaeum.
rektum, utsöndringskanaler och genitala kanaler öppnar sig i cloaca, och det öppnar sig till utsidan med en kloaköppning. Men i många Benfiskar och alla däggdjur (utom monotremer) har rektum och urinogenitala kanaler separata öppningar till utsidan; öppningen av den förra är en anus.
rektum hos däggdjur är inte homologt med rektum hos ryggradsdjur eftersom det hos däggdjur härrör från uppdelning av embryonal cloaca. I de flesta ryggradsembryon finns en postanal tarm som en förlängning av tarmen i svansen, men den försvinner senare.
i elasmobranchs bär tjocktarmen ett par rektala körtlar som utsöndrar slem och natriumklorid. I amnioter finns en ileokolisk ventil vid korsningen av små och stora tarmar, som saknas i fiskar. Det förhindrar bakterier att komma in i ileum från kolon.
i amnioter från denna korsning uppstår en ileocolic caecum som är två i fåglar. Den innehåller cellulosa som smälter bakterier. Det är mycket länge i växtätande däggdjur (kanin, häst, ko, etc.). I primater är caecum liten med en vestigial vermiform bilaga.
matsmältningskörtlar:
1. Lever:
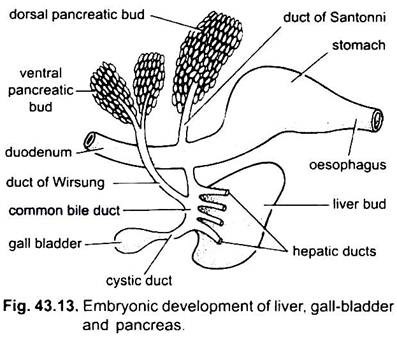
levern uppstår som en enkel eller dubbel utväxt från den ventrala väggen i den embryonala archenteronen. Denna utväxt bildar en ihålig leverdivertikulum, som snart skiljer sig åt i en främre del, som prolifererar för att bli levern och dess gallkanaler, och en bakre del, vilket ger upphov till gallblåsan och cystisk kanal. Gallgångarna förenas för att bilda en leverkanal som förenas med den cystiska kanalen för att bilda en vanlig gallkanal eller ductus choledochus. Regionen av archenteron från vilken levern uppstår blir duodenum.
levern är den största lobade körteln i kroppen, suspenderad av ett dubbelt lager av bukhinnan från tvärgående septum eller dess representant.
en gallblåsa är för lagring av galla som utsöndras av levercellerna, ligger i levern och dränerar in i tolvfingertarmen genom vanlig gallkanal som bildas av föreningen av cystisk kanal och leverkanal. En gallblåsa är inte oumbärlig och saknas hos många fåglar och däggdjur.
en lever finns i alla ryggradsdjur. I cyklostomer är den liten, singelflikad (lampreys) och två flikad i hagfishes. Det är bilobed i elasmobranchs, två eller tre lobed i Benfiskar, amfibier, reptiler och fåglar och många lobed i däggdjur. Leveren är lång, smal och cylindrisk i fiskar, urodeler och ormar.
den är kort, bred och platt i fåglar och däggdjur. En gallblåsa och gallkanal finns i larvcyklostomer men de är frånvarande hos vuxna. Fiskar, amfibier och reptiler har i allmänhet en gallblåsa, men det saknas hos många fåglar. De flesta däggdjur har en gallblåsa, men det är frånvarande i Cetacea och Ungulata.
levern utsöndrar en vattnig, alkalisk galla men har inga enzymer. Det neutraliserar surheten hos mat som kommer in i tolvfingertarmen. Det hjälper i matsmältningen av fetter.
2. Bukspottkörteln:
bukspottkörteln bildas från endoderm av den embryonala archenteronen. En enda dorsal divertikulum från embryonal duodenum och en eller två ventrala utväxter från levern bildar bukspottskörteldivertikula. De proximala delarna av divertikula bildar bukspottkörtelkanaler, men dessa kanaler genomgår reduktion eller fusion så att endast en eller två bukspottkörtelkanaler förblir hos vuxna, de öppnar sig i duodenum antingen separat eller efter att ha förenats med den gemensamma gallkanalen.
de distala delarna av divertikula genomgår spirande för att bilda huvudmassan av bukspottkörtelceller till vilka mesodermala derivat tillsätts. Således görs en enda körtel som har flera lober som bildar antingen en diffus eller en kompakt bukspottkörtel.
bukspottkörteln är både en exokrin och endokrin körtel, bunden av känsliga bindvävsträngar. Den exokrina delen utsöndrar matsmältningsenzymer som hälls i duodenum genom bukspottkörtelkanaler. Medan den endokrina delen utsöndrar hormoner som insulin och glukagon.
en bukspottkörtel finns i alla ryggradsdjur. I lampreys, några beniga fiskar, lungfiskar och lägre tetrapoder, är det ett diffust organ inbäddat i levern, mesenterierna och tarmväggen. Hagfishes har en liten bukspottkörtel. Elasmobranchs har en väldefinierad bilobed bukspottkörtel. I högre tetrapoda är det i allmänhet en kompakt körtel. En eller två bukspottkörtelkanaler öppnar sig i tolvfingertarmen.