- introduktion
- resultat
- DBH-Cre-drivrutinen markerar och är begränsad till th-Uttryckande noradrenerga regioner i hjärnstammen som hämmas av CNO-administrering
- CNO-hM4D-medierad störning av noradrenerga neuroner hos vuxna möss
- CNO-hM4D-störning av DBH-Cre-neuroner resulterar i ett Hyperkapniskt underskott
- Habituerade Syskonkontroller injicerade med 10 mg / kg CNO visade ett Hyperkapniskt Ventilationsunderskott
- Habituerade och Ohabituerade Vildtypsdjur injicerade med saltlösning visade inga förändringar Presaline och Postsaline
- Klozapin till CNO-Koncentrationsförhållandena är högre i hjärnan än i Serum
- diskussion
- material och metoder
- etiskt godkännande
- avel, genetisk bakgrund och underhåll av möss
- histologi
- elektrofysiologi
- pletysmografi
- vätskekromatografi-masspektrometri
- datainsamling och analys
- pletysmografi
- statistik
- pletysmografi
- vätskekromatografi-masspektrometri
- Etikuttalande
- Författarbidrag
- finansiering
- intressekonflikt uttalande
- bekräftelser
introduktion
de neurala nätverk som ligger till grund för respiratorisk kemosensorisk reflex är ett primärt mål för att förstå etiologin för flera beteendemässiga och fysiologiska störningar. Störda kemosensoriska reflexer antas spela en roll i både medfödda och vuxna störningar inklusive plötsligt spädbarnsdödssyndrom (SIDS), medfödd Central Hypoventilationssyndrom (CCHS), central sömnapneas, onormal andning i Rett syndrom och fetma hypoventilationssyndrom (Obonai et al., 1998; Ozawa et al., 2003; Amiel et al., 2009; Ramanantsoa et al., 2011; Lavezzi et al., 2013). Under den falska kvävningslarmhypotesen tros olämplig kemosensorisk aktivering eller överkänslighet spela en roll i undergrupper av patienter som lider av panikstörning (Klein, 1993). Kemosensorisk dysfunktion kan också spela en roll i neurodegenerativa sjukdomar genom sömnstörande andning som är associerad med accelererad progression (Hakim et al., 2012; Verbraecken och McNicholas, 2013; Bahia och Pereira, 2015; Snyder et al., 2017). Således kommer en bättre förståelse av hjärnstammens kemosensoriska nätverk att ge viktiga ledtrådar till ett antal beteendemässiga och fysiologiska patologier.
Farmakogenetiska designerreceptorer aktiveras exklusivt av designerläkemedel (DREADDs) (Armbruster et al., 2007) har använts i ett antal studier för att kartlägga neurala populationer i respiratorisk kemoreflex (andningsrespons på förhöjda CO2-nivåer i blodet). DREADD-teknik i kombination med intersektionell genetisk utplacering har använts för att tysta mycket riktade neuronpopulationer medan man observerar andningsfunktionen i medvetna och obegränsade möss av vårt laboratorium och andra, vilket undviker många förvirringar från tidigare kretsmappningsmetoder (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sol och stråle, 2017; sol et al., 2017). Dessa och de flesta andra studier inkluderade nästan alltid CNO endast icke-DREADD-Uttryckande syskonkontroller som inte visade några kemosensoriska eller andra andningseffekter och hävdade att CNO inte hade några off-target-effekter på andning i medvetna och obehindrade möss trots den höga dosen som användes, 10 mg/kg. Ändå hittades det i flera andra studier att CNO och dess ryggmetabolismsprodukter kunde ha off-target effekter på beteende och rörelse i en mängd olika analyser, men andningsutgången behandlades inte (Guettier et al., 2009; Joober, 2010; MacLaren et al., 2016; Gomez et al., 2017; Ilg et al., 2018; Mahler och Aston-Jones, 2018; Manvich et al., 2018; Padovan-Hernandez och Knackstedt, 2018). Det visades också att CNO och dess metaboliter inte är likvärdigt fördelade mellan cirkulationssystemet och hjärnan (Gomez et al., 2017).
en andra, oadresserad oro uppstår från den stress som induceras av det experimentella paradigmet, inklusive att hålla djur i en barometrisk pletysmografikammare för hela kroppen, hantering, intra-peritoneal injektion och rektala temperaturmätningar. I tidigare DREADD-respiratoriska studier infördes na-mice-möss i en liten kammare (140-400 mls) och fick acklimatisera 20-40 min före datainsamling (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sol och stråle, 2017; sol et al., 2017). Det är emellertid inte klart om denna tid är tillräcklig för att minimera stressinducerade andningsförändringar som kan fungera som en interagerande faktor med effekterna av clozapin.
i våra studier för att undersöka det noradrenerga (NA) systemets roll i andningskontrollen försökte vi använda den väletablerade RC::P_hM4D DREADD allel (Ray et al., 2011) för att testa rollen för NA-neuroner i baslinjen och hyperkapnisk andning hos ohämmade vuxna djur med helkroppspletysmografi. Eftersom noradrenalin och NA-neuroner också är kända för att spela en central roll i stressresponser (Valentino och Van Bockstaele, 2008; Chen et al., 2018) genomförde vi en serie studier för att jämföra omfattande tillvänjning (flera exponeringar mot det experimentella paradigmet där djuret lär sig att upplevelsen inte är hotande eller överlevande) och hög vs. låga (1 mg/kg) CNO-doser med tidigare publicerade DREADD-protokoll (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sol och stråle, 2017; sol et al., 2017). Här visar vi för första gången att höga systemiska doser av CNO kan framkalla off-target-effekter på en autonom andningsfunktion hos medvetna möss. Vi avslöjar också att off-target-effekten av CNO på kemosensorisk andningsutgång effektivt avslöjas av omfattande tillvänjning och skulle därför inte vara uppenbart i tidigare CNO-kontrollstudier som inte hade vana djur före andningsmätning, med endast en kort acklimatiseringsperiod före datainsamling. Tillsammans tyder dessa resultat på att tidigare kartlagda neuronpopulationer indirekt kan påverka andningskontrollen genom potentiella roller vid reglering av stressresponser. Dessa data stämmer särskilt överens med de senaste rapporterna som tyder på att CNO inte är biologiskt inert vid höga doser via metabolisk omvandling till klozapin (MacLaren et al., 2016; Gomez et al., 2017) och att beteendeeffekter utanför målet kan manifestera sig inte bara från störning av beteendekretsar utan också från störningar till underliggande autonoma kretsar och homeostas.
resultat
DBH-Cre-drivrutinen markerar och är begränsad till th-Uttryckande noradrenerga regioner i hjärnstammen som hämmas av CNO-administrering
för att undersöka uttrycket och specificiteten för DBH-Cre-linjen använde vi ett enda rekombinasavelsschema (Figur 1a) där vi korsade föraren till Ai9-linjen (Madisen et al., 2010), som uttrycker en floxed tdTomato. Färgning med en tyrosinhydroxylas (TH) antikropp avslöjade att uttryck av tdTomato i hjärnstammen var begränsad till th-Uttryckande regioner, inklusive locus coeruleus, A5, A1, A2, A7, SubCV och SubCD-kärnor som förväntat (Figur 1b). För att bekräfta att NA-neuroner som uttryckte hm4d-receptorn var lyhörda för CNO, utförde vi inspelningar på locus coeruleus (LC) neuroner, där vi observerade en hämning av neuronbränning vid CNO-badapplikation (n = 3, Figur 1C).
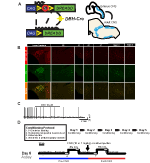
Figur 1. Avel schematiska och respiratoriska protokoll. RC::P_hM4D-allel kombineras med en dopamin-beta-hydroxylas (DBH)-Cre-linje för att uppnå cellspecifikt uttryck av hM4D-receptorn i noradrenerga (NA) neuroner (a). Fluorescerande uttryck av tdTomato i DBH-Cre; Ai9 koronala hjärnsektioner samuttrycker med tyrosin-hydroxylas (TH) i hjärnstammens NA-kärnor (B). I skiva inspelning av NA locus coeruleus före och efter CNO ansökan, visar DREADD receptor funktionalitet i riktade neuroner (C). Andningsprotokoll. Habituerade möss utsätts för ett 5-dagars habituerat protokoll bestående av hantering, mätning av rektal sondtemperatur, saltinjektion och exponering för pletysmografikammaren. På den sjätte dagen, både vana och ohabituerade möss under en hyperkapnisk analys där djuret placeras i andningskammaren och får acklimatisera sig under rumsluftförhållanden vid baslinjen. Djuret exponeras sedan för 20 min 5% CO2 följt av 20 min rumsluft. Djuret injiceras sedan intraperitonealt med CNO eller saltlösning, följt av ytterligare 20 min rumsluft efter injektion, 20 min 5% CO2 och 20 min rumsluft. Öppna rutor avgränsa datainsamlingstider (D).
CNO-hM4D-medierad störning av noradrenerga neuroner hos vuxna möss
för att undersöka NA-neurons roll under baslinjen och hyperkapnisk andning använde vi RC:P_hM4D-hämmande DREADD-system korsat med DBH-Cre-föraren. Använda helkroppspletysmografi (Ray et al., 2011) mätte vi ventilationssvaren hos ohämmade vuxna djur under rumsluft (21% O2/79% N2) och hyperkapnisk (5% CO2/21% O2/74% N2) förhållanden före och efter CNO-administrering (figur 1D). För att ta itu med CNO-dosering och potentiell stress inducerad av vår experimentella design utsattes djur för ett av fyra tillstånd: (1) unhabituated och injicerades med 10 mg/kg CNO; (2) habituated och injicerades med 10 mg/kg CNO; (3) unhabituated och injicerades med 1 mg/kg CNO; eller (4) habituated och injicerades med 1 mg/kg CNO. Tillvänjning bestod av en 5-dagars process som innebar hantering, rektal temperatursond, saltinjektion och exponering för pletysmografikammaren varje dag i 30 minuter, medan na-djur endast exponerades för en 20-45 min kammaracclimationsperiod omedelbart före datainsamling som gjort i tidigare studier (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sol och stråle, 2017; sol et al., 2017). Respiratoriska parametrar uppmätta inkluderade andningsfrekvens RR, tidvattenvolym (VT), minutventilation (VE), syreförbrukning (VO2), minutventilation normaliserad till syreförbrukning (VE/VO2), apnea-frekvens, suckfrekvens och variationskoefficienter för interbreath intervall och amplitud (periodisk och volyminstabilitet). Som en ytterligare kontroll jämförde vi också vana och ohabituerade vildtypsdjur injicerade med saltlösning.
CNO-hM4D-störning av DBH-Cre-neuroner resulterar i ett Hyperkapniskt underskott
under tre av förhållandena, DBH-Cre; RC::P_hm4d-djur visade en minskning av VE Och VE / VO2 efter CNO-administrering, medan syskonkontroller inte visade några skillnader i pre-och post-CNO-värden. Ohabituerade djur injicerade med 10 mg/kg CNO visade en signifikant reducerad RR (-12, 17%, p = 0, 034) och VT (-30, 87%, p = 0, 0016), vilket resulterade i en minskning av VE (-38, 64%, p = 0, 0031) och en liten minskning av VO2 (-14, 25%, p = 0, 042) (Figur 2). Minskningen I VE var större än minskningen i VO2, vilket resulterade i en total minskning av VE/VO2 (-26, 89%, p = 0, 0095). Ohabituerade djur injicerade med 1 mg/kg CNO visade en trend mot minskad RR (-12,88%, p = 0,066) och signifikant reducerad VT (-16,52%, p = 0,00085) och VE (-28,08%, p = 0,0070), vilket ledde till en total minskning av VE/VO2 (-22,23%, p = 0,016) (Figur 3). Slutligen visade vana djur injicerade med 1 mg/kg CNO en reducerad RR (-10,77%, p = 0,074), trend mot reducerad VE (-25,07%, p = 0,074) och minskad total VE/VO2 (-23,70%, p = 0,024) (Figur 4). Inga signifikanta förändringar i apnea-frekvens, suckfrekvens, periodisk eller volyminstabilitet eller temperatur sågs i någon kohort.
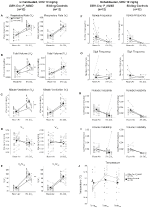
Figur 2. Unhabituated DBH-Cre; RC:: P_hM4D djur administrerade 10 mg/kg CNO visar ett hyperkapniskt underskott medan syskonkontroller inte visar någon förändring. Efter CNO administration, DBH-Cre; RC::P_hm4d-djur visar ingen förändring i rumsluftventilation och minskningar av RR (a), VT (B), VE (C), VO2 (D) och VE/VO2 (E) under hyperkapniska förhållanden utan förändringar i apnea-frekvens (F), suckfrekvens (G), periodisk (H) eller volyminstabilitet (I) och temperatur (J). Syskonkontroller visade ingen skillnad i alla pre – och post-CNO-värden.
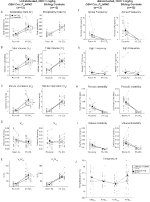
Figur 3. Unhabituated DBH-Cre; RC:: P_hM4D djur administrerade 1 mg/kg CNO visar ett hyperkapniskt underskott medan syskonkontroller inte visar någon förändring. Efter CNO administration, DBH-Cre; RC::P_hm4d-djur visade ingen signifikant förändring i rumsluftventilation eller RR (a) och visade minskningar av VT (B), VE (C) och VE/VO2 (E) under hyperkapniska förhållanden utan förändringar i VO2 (D), apnea-frekvens (F), suckfrekvens (G), periodisk (H) eller volyminstabilitet (I) och temperatur (J). Syskonkontroller visade ingen skillnad i alla pre – och post-CNO-värden.
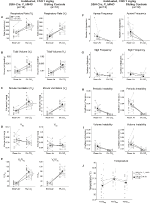
Figur 4. Habituated DBH-Cre; RC:: P_hM4D djur administrerade 1 mg/kg CNO visar ett hyperkapniskt underskott medan syskonkontroller inte visar någon förändring. Efter administrering av CNO, DBH-Cre; RC:: p_hm4d djur visar ingen förändring i rumsluftventilation och en minskning av RR (A) och VE/VO2 (E) under hyperkapniska förhållanden utan några signifikanta förändringar i VT (B), VE (C), VO2 (D), apnea-frekvens (F), suckfrekvens (G), periodisk (H) eller volyminstabilitet (I) och temperatur (J). Syskonkontroller visade ingen skillnad i alla pre – och post-CNO-värden.
Habituerade Syskonkontroller injicerade med 10 mg / kg CNO visade ett Hyperkapniskt Ventilationsunderskott
i båda DBH-Cre; RC::P_hm4d och syskonkontroll habituerade djur injicerade med 10 mg / kg CNO, vi noterade en signifikant minskning av VE / VO2 (p = 0.0235) medierad av minskningar i RR (p = 0.00036) och VE (p = 0.037) (Figur 5). Till skillnad från de andra kohorterna var det emellertid ingen skillnad mellan DBH-Cre; RC::P_hM4D och syskonkontrolldjur i dessa parametrar: RR (-12,72 vs. -10,22%, p = 0,6268), VE (-24,88 vs. -23,94%, p = 0,4150) eller VE/VO2 (-15,4 vs. -22,55%, p = 0,4643).
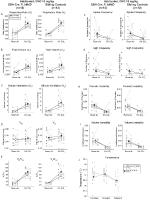
Figur 5. Båda vana DBH-Cre; RC::P_hm4d djur och syskonkontroller administrerade 10 mg / kg CNO visar ett hyperkapniskt underskott. Efter CNO-administrering visar DBH-Cre; RC::p_hm4d djur och syskonkontroller ingen förändring i rumsluftventilation och en minskning av RR (a), VE (C) och VE/VO2 (E) under hyperkapniska förhållanden utan signifikanta förändringar i VT (B), VO2 (D), apnea-frekvens (F), suckfrekvens (G), periodisk (H) eller volyminstabilitet (I) och temperatur (J).
Habituerade och Ohabituerade Vildtypsdjur injicerade med saltlösning visade inga förändringar Presaline och Postsaline
även om inga fenotyper observerades i kontroller injicerade med 1 mg/kg CNO, behandlade vi möjligheten att själva injektionen orsakade fenotypen observerad i kontroller injicerade med 10 mg/kg CNO genom att testa habituerade och ohabituerade vildtypsdjur injicerade med saltlösning (Figur 6). I både vana och ohabituerade kohorter visade djur ingen skillnad i respiratoriska parametrar presalinadministration och postsalinadministrering. Inga signifikanta förändringar i apnea-frekvens, suckfrekvens, periodisk eller volyminstabilitet eller temperatur sågs.
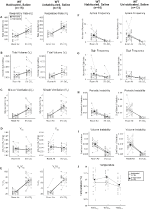
Figur 6. Både ohabituerade och vana vildtypsdjur administrerade saltlösning visar ingen skillnad mellan värden före och efter injektion. Efter administrering av saltlösning visade både ohabituerade och vana vildtypsdjur inga förändringar under rumsluft eller hyperkapniska förhållanden i RR (a), VT (B), VE (C), VO2 (D), VE/VO2 (E), apnea-frekvens (F), suckfrekvens (G), periodisk (H) eller volyminstabilitet (I) och temperatur (J).
Klozapin till CNO-Koncentrationsförhållandena är högre i hjärnan än i Serum
för att bestämma biotillgängligheten för CNO och klozapin mätte vi deras koncentrationer i serum och hjärna via masspektrometri. Trettio minuter efter en intraperitoneal injektion av CNO hos möss finns CNO i lägre relativ överflöd jämfört med dess bakmetabolit clozapin i serum och hjärna för alla testade doser, 0, 1 mg/kg (serum p = 0, 0054, hjärna p = 0, 0001) (figur 7A), 1 mg/kg (serum p > 0, 05, hjärna p = 0, 0197) (figur 7B) och 10 mg/kg (serum p = 0, 0036, hjärna p = 0.0005) (figur 7C). När det totala förhållandet mellan klozapin och CNO analyserades var de alltid över noll och var högre i hjärnan än i serum (vehikel p > 0, 5, 0, 1 mg/kg p > 0, 5, 1 mg/kg p = 0, 0018, 10 mg/kg p = 0, 0160) (figur 7D).
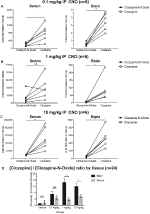
Figur 7. Klozapin till CNO-koncentrationsförhållandena är högre i hjärnan än i serum. Serum-och hjärnprover samlades in från vildtypsmöss 30 min efter CNO-administrering i 3 olika doser 0,1 mg/kg (a), 1 mg/kg (B), 10 mg/kg (C). Individuella datapunkter representerar det relativa överflödet av klozapin-N-oxid eller klozapin erhållet med LC-MS. Klozapin till CNO-förhållande för varje dos och SEM visas också (D).
diskussion
det ursprungliga målet med denna studie var att undersöka rollen av DBH-Cre-neuroner i andningsfysiologi efter akut störning hos obehindrade och medvetna vuxna djur. Eftersom noradrenalin och NA-neuroner spelar en väldokumenterad roll i stressbeteenden (Valentino och Van Bockstaele, 2008; Chen et al., 2018) försökte vi också undersöka om tillvänjning till ett sannolikt stressande fysiologiskt protokoll skulle ha en effekt på de respiratoriska fenotyperna som tidigare observerats. I hela kroppen pletysmografi DREADD-protokollet som används i vårt laboratorium och andra hanteras djur, rektalt probed för temperatur, utsätts för en ny miljö (pletysmografikammaren) och injiceras intraperitonealt. Tidigare studier har visat att hantering och tillvänjning till injektion och andra ”rutinmässiga” procedurer kan modifiera beteendemässiga och fysiologiska parametrar, inklusive andning (Misslin et al., 1982; Andrews och fil, 1993; Lapin, 1995; Ryabinin et al., 1999). Andra tillämpade stressorer modifierar också andning under både baslinje-och hyperkapniska ventilationsförhållanden (Isom och Elshowihy, 1982; Kinkead et al., 2001).
i våra studier med höga doser av CNO fann vi att hM4D-medierad hämning av DBH-Cre-definierade NA-neuroner resulterade i en reducerad hyperkapnisk reflex I VE/VO2 i fyra definierade experimentella kohorter, med reducerad RR, VT och VE, som stöder tidigare studier (Biancardi et al., 2008; Viemari, 2008; Gargaglioni et al., 2010). Emellertid visade habituerade syskonkontroller med en dos på 10 mg/kg CNO som användes i tidigare andningsstudier ett ventilationsunderskott under hyperkapniska förhållanden av samma storlek som det som ses i DBH-Cre; RC::P_hM4D djur. Inga andra syskonkontrollgrupper visade denna fenotyp, inklusive den vana kohorten som endast fick (1 mg/kg CNO) eller saltlösning. Dessa resultat tyder på att högre CNO-doser (10 mg/kg) har en effekt på det hyperkapniska svaret som avslöjas efter omfattande tillvänjning samtidigt som det förmodligen minskar djurstressnivåerna, och att lägre CNO-doser inte har någon effekt på andningskontrollen hos Vana djur. Dessa resultat överensstämmer också med arbete från Korsak et al. who visade att lågdos CNO (2 mg/kg) inte ger effekter utanför målet i arbetsförmåga i en träningsanalys som inkluderade tidigare träning (Korsak et al., 2018) och Fleury Curado et al. who visade inga låga doser CNO (1 mg/kg) specifika effekter på andningsutgången (Fleury Curado et al., 2018).
de ökade förhållandena mellan klozapin och CNO-nivåer i serum och i hjärnan (Figur 7) överensstämmer med nya studier som tyder på att CNO lätt metaboliseras tillbaka till klozapin och visar större hjärnpermeabilitet jämfört med CNO hos möss och på andra håll (Jann et al., 1994; Chang et al., 1998; Guettier et al., 2009; Gomez et al., 2017; Raper et al., 2017). Det är emellertid inte klart om de observerade off-target-effekterna beror på CNO eller clozapin. Eftersom våra hyperkapniska mätningar inträffade < 30 minuter efter CNO-applicering är det troligt att andningseffekterna utanför målet medieras av clozapin. Våra resultat (Figur 7) visar höga relativa nivåer av klozapin i hjärnan, även om CNO inte är helt frånvarande. Huckstepp och kollegor använde emellertid direkt CNO-applikation på ventralmedulla hos bedövade råttor för att visa att endast vid rumsluft och inte under hyperkapniska eller hypoxiska utmaningar har CNO-applikation en liten effekt, ökande frekvens och minskande expiratorisk varaktighet men lämnar VT oförändrad, utan någon tydlig effekt observerad under hyperkapni (Huckstepp et al., 2015). Med tanke på den direkta appliceringen i bedövade råttor och tidsramen för experiment är det troligt att de små off-target-effekterna som ses medierades av CNO och inte av clozapin.
den bakre metaboliten clozapin är ett vanligt förekommande lugnande och antipsykotiskt läkemedel vid schizofreni med många endogena mål, inklusive antagonistiska åtgärder med låg hög affinitet vid D1 -, D2-och D4-dopaminerga receptorer, 5-HT2A, 5-HT2C, 5-HT3, 5-HT6 och 5-HT7 serotonerga receptorer, H1-histaminreceptorer, och bland annat adrenerga receptorer för adrenerga receptorer (Fitton och Heel, 1990; Ashby och Wang, 1996). De off-target-effekter som ses här kan bero på distinkta eller kombinerade mekanismer och mål. Klozapin kan påverka andningen som ett lugnande medel. En tidigare studie visade minskad RR och VT under 5 och 10% CO2 under både slow-wave-sleep och rapid-eye-movement sömntillstånd jämfört med det tysta vakna tillståndet hos möss (Nakamura et al., 2007). Alternativt kan hämning av riktade DREADD-Uttryckande neuroner resultera i en antianxiogen eller anxiolytisk effekt som liknar vårt habituationsprotokoll för att avslöja CNO/clozapinmedierade kemosensoriska off-target-effekter. Båda förklaringarna stöds av flera studier som har visat att CO2 spelar en roll i medfödda och lärda rädslesvar och ångestrelaterade beteenden (Ziemann et al., 2009; Feinstein et al., 2013; Taugher et al., 2014; Dlouhy et al., 2015; vinter et al., 2017). Således kan neuroner riktade i några av dessa studier verkligen spela en roll för att driva anxiogena beteendemässiga svar snarare än fysiologiska kemosensoriska reflexer, eftersom både katekolaminerga och serotonerga system är involverade i rädsla/ångestbeteenden och kemosensorisk homeostas (Brust et al., 2014; Hennessy et al., 2017).
omvänt kan de kemosensoriska fenotyperna som observerats med höga CNO-nivåer vara äkta, eftersom vi kunde rekapitulera det NA-medierade hyperkapniska underskottet vid CNO-doser, en storleksordning lägre i vana möss, medan kontrollgrupper inte visade någon CNO-effekt. Fullständiga jämförelser mellan tidigare studier är dock svåra på grund av bristen, i vissa fall, av rapporterade VO2 -, VT -, RR-och VE/VO2-data. Till exempel kan förändringar i kroppstemperatur eller metabolisk hastighet också påverka på ett antal sätt på andnings-och kemosensorisk utgång, och att pletysmografkammartemperaturerna var väldigt olika i vissa fall (34 mot 30 kcal C i våra studier), vilket påverkar det dynamiska området för den barometriska komponenten i vågformen och därmed tidvattenvolymen. I synnerhet såg vi varken märkbara förändringar i andningsvågformsegenskaper i något av våra tillstånd eller akut hjärt-andningsstopp som observerades i våra tidigare höga doser, hela rombomerstudier (Sun et al., 2017).
våra resultat visar för första gången att CNO har en oväntad effekt på den hyperkapniska kemosensoriska reflexen som avslöjas av omfattande tillvänjning. Viktigt, trots höga nivåer, hade en CNO-off-target-effekt tidigare uteslutits på grund av bristen på fenotyp i syskonkontroller, men som vi visar blir tydliga vid tillvänjning. Vi erbjuder en off-target karakterisering av CNO i musmodellsystemet för att komplettera studier på råtta och icke-mänskliga primater. Resultaten här höjer möjligheten att ytterligare CNO-medierade, off-target-effekter på kretsarna som studeras eller till autonoma eller homeostatiska kretsar kan existera men kan maskeras i andra kontrollerade experiment. Viktigt är att dessa data avslöjar att utredare bör sträva efter att använda de minimala doserna av den aktiverande liganden som är möjliga i kombination med höga nivåer av tillvänjning, och att de korrekta kontrollerna måste ingå i kemiska genetiska manipuleringar för att fullt ut uppskatta och tolka experimentella data.
material och metoder
etiskt godkännande
studier godkändes av Baylor College of Medicine Institutional Animal Care and Use Committee enligt protokoll AN-6171.
avel, genetisk bakgrund och underhåll av möss
vi upprätthöll kolonier av alla våra heterozygota musstammar genom backcrossing till C57BL/6J Möss och homozygota stammar av syskonkorsningar. För histologiska experiment parades DBH-Cremice med den homozygota AI9-musen (Madisen et al., 2010) (JAX 007909). För pletysmografiexperiment parades DBH-Cre-möss med homozygot RC:: P_hM4D (Ray et al., 2011) möss för att härleda djur, där alla möss Bar RC::P_hM4D-allelen. Syskondjur som inte ärvde Cre-allelen användes som kontrolldjur till Cre-positiva avkommor. Rosa26 – specifika primrar för Ai9, RC::P_hM4D och RC:: Epe-möss var 5′-GCACTTGCTCTCCCAAAGTC, 5′-GGGCGTACTTGGCATATGAT och 5′ – CTTTAAGCCTGCCCAGAAGA och gav ett 495 bp-band (riktat) och 330 bp-band (wt). Cre-specifika primers för alla rhombomere Cre-drivrutiner var 5 ’- ATCGCCATCTTCCAGCGCGCACCATTGCCC och 5’-GCATTTTCTGGGGGATTGCTTA och gav ett 550 bp-band om det var positivt. För LC-MS-experiment erhölls C57BL / 6J-möss från Center of Comparative Medicine (CCM), Baylor College of Medicine.
histologi
fyra till åtta veckor gamla DBH-Cre; Ai9 vuxna möss perfunderades transkardiellt med 0,1 M fosfatbuffrad saltlösning (PBS) och sedan med 4% paraformaldehyd (PFA) i PBS. Hjärnor dissekerades ut och föll fixerade för 2 h i 4% PFA före en PBS-sköljning och jämvikt i 20% sackaros i PBS. Hjärnor sektionerades sedan in i 30 occurm-sektioner, monterade på glidbanor och märktes med immunofluorescerande antikroppar. Vi färgade över natten med anti-tyrosinhydroxylasantikropp för att identifiera katekolaminerga neuroner (1:1,000, Millipore AB152) följt av 2 h med donkey Anti-rabbit Cy3 sekundär (1: 500, Jackson 711-165-152) i 0.1% Triton-X i PBS (PBST) med 5% donkey serum. Bilder samlades på ett Zeiss upprätt epifluorescerande mikroskop.
elektrofysiologi
horisontella hjärnskivor innehållande locus coeruleus (300 oc.m) skars med en vibratom (Leica VT 1000s, Leica Microsystems, Buffalo Grove, IL) från vuxen DBH-Cre; RR2P; RC::Epe-möss (~1 månad gammal) i 4 c c artificiell cerebrospinalvätska (ACSF). Skivorna nedsänktes i en kammare perfuserad med syresatt ACSF (95% O2 och 5% CO2) innehållande i mM: 124 NaCl, 2,0 KCl, 1,3 MgSO4, 2,5 CaCl2, 1,2 KH2PO4, 25 NaHCO3 och 10 glukos (2-3 ml/min). Hela cellinspelningar utfördes vid 30 2CB C med användning av konventionella patch-clamp tekniker och en MultiClamp 700B förstärkare (Molecular Devices, Union City, CA). GFP-positiva neuroner från locus coeruleus identifierades visuellt och avbildades därefter med infraröd differentialinterferenskontrastvideo på scenen i ett upprätt mikroskop (Axioskope FS2, Carl Zeiss, Oberkochen, Tyskland). Patchpipetter (resistanser 4-6 MU) fylldes med (i mM): 110 K-glukonat, 10 KCI, 10 HEPES, 10 Na2-fosfokreatin, 2 Mg3-ATP och 0,2 Na3-GTP; pH justerades till 7,2 och osmolaritet till 300 mOsm. Innehavspotentialen var -70 mV. CNO var bad tillämpas.
pletysmografi
pletysmografi på medvetna, ohämmade möss utfördes som beskrivs på 6 – till 12 veckor gamla vuxna djur (Ray et al., 2011). Habituerade möss utsattes för ett 5-dagars habituationsprotokoll med varje dag bestående av flera minuters hantering, temperatur tagen av rektal sond, intraperitoneal saltinjektion och 30 min i pletysmografikammaren. Möss testades sedan inom 1 vecka efter den sista dagen av tillvänjning. Ohabituerade möss utsattes inte för hantering eller pletysmografkammaren. Alla möss var na ouguive till CNO och används endast en gång.
på testdagen togs möss från sin hembur, vägdes och rektaltemperaturen togs. Djur placerades sedan i en genomströmning, temperaturkontrollerad (vattenmantlad vid 30 CCB) pletysmografikammare och fick acklimatisera sig i minst 20 minuter i rumsluft (21% O2/79% N2) förhållanden. Efter acklimatisering och mätning under rumsluft byttes kammargasen till en hyperkapnisk blandning av 5% CO2/21% O2/74% N2 under 20 min. Kammargasen växlades sedan tillbaka till rumsluften i 20 minuter. Mössen avlägsnades kort för rektal temperaturmätning och intra-peritoneal injektion av CNO (National Institute of Mental Health Chemical Synthesis and Drug Supply Program) upplöst i saltlösning (1 eller 0,1 mg/ml) för en effektiv koncentration av 10 respektive 1 mg/kg. Djuret återlämnades till kammaren för ytterligare 20 min rumsluft, 20 min hyperkapnia och 20 min rumsluft. Djuret avlägsnades sedan från kammaren och rektaltemperaturen togs omedelbart efter avslutad experiment.
vätskekromatografi-masspektrometri
tjugofyra vildtypsmöss, jämnt uppdelade efter kön, vägdes och behandlades med 10 mg/kg CNO, 1 mg/kg CNO, 0,1 mg/kg CNO eller vehikel. Trettio minuter efter injektion samlades blodprover via hjärtpunktur och placerades i BD-Mikrobehållare. Proverna centrifugerades vid 4 kg C vid 13 500 rpm i bänkcentrifug och supernatanter samlades in. Serum och hjärnor hölls vid -20 kcal C tills extraktion.
HPLC-lösningsmedel vatten, acetonitrilkloroform och metanol och masspektrometri-myrsyra erhölls från Sigma-Aldrich (St.Louis, MO). Kalibreringslösning innehållande flera kalibreringsmedel i en lösning av acetonitril, trifluorättiksyra och vatten köptes från Agilent Technologies (Santa Cruz, CA). Metaboliter och interna standarder, inklusive N-acetyl asparaginsyra-d3, tryptofan-15n2, glutaminsyra-d5, tymin-d4, gibberellinsyra, trans-zeatin, jasmonsyra, 15N antranilsyra och testosteron-D3 köptes från Sigma-Aldrich (St.Louis, MO). Microtainer R SST TM erhölls från Becton Dickinson (Franklin Lakes, NJ).
extraktion bestod av 750 occyl iskall metanol:vatten (4:1) innehållande 20 occyl spikade interna standarder som tillsattes till varje hjärnprov (50 mg) och kvalitetskontroller och sedan homogeniserades i 1 min vardera. Sedan tillsattes 750 UCL av 100% acetonitril innehållande 20 UCL spikade interna standarder för att tvätta provet (100 UCL) och kvalitetskontroller och sedan sonicated för 5 min. Alla prover centrifugerades vid 5000 rpm för 10 min vid 4 kcal C. Den resulterande supernatanten samlades in, och 20 oC-l injicerades i LC-MS.
alla prover analyserades med användning av 6,490 trippel quadrupol masspektrometer (Agilent Technologies, Santa Clara, CA) kopplad till HPLC-system (Agilent Technologies, Santa Clara, CA) genom multipel reaktionsövervakning (MRM). Cirka 8-11 datapunkter förvärvades per detekterad metabolit. Detekterade metaboliter var klozapin, CNO och norklozapin (N-Desmetylklozapin). ESI-positivt läge användes i metoden. HPLC-kolonnen var ACQUITYUPLC C18-kolonn (100 kcal, 1,8 kg och 2,1 mm 100 mm. Milford, MA, USA) med en flödeshastighet på 0,5 ml/min.
datainsamling och analys
pletysmografi
pletysmografi tryckförändringar mättes med användning av en Validyne dp45 differentialtrycksgivare och referenskammare och CD15 bärare demodulator och registreras med LabChartPro i realtid. Vågformer analyserades offline med hjälp av LabChartPro och anpassad skriftlig MATLAB-kod för att bestämma andningsfrekvens (RR), tidvattenvolym (VT) (Ray et al., 2011), minutventilation (VE) och mönsteranalys. Andningsvågformer samlades offline under perioder då djuret var i vila och avläsningar var fria från rörelseartefakter. Minst 1 min kumulativa data som sammanställts från spår som är minst 10 s långa från den sista 10 min av ett givet experimentellt tillstånd analyserades. O2-förbrukningen bestämdes genom att jämföra gaskompositionen mellan kalibrering i en tom kammare och levande andning med hjälp av en AEI-syresensor och analysator. Kammartemperaturen övervakades ständigt med hjälp av en ThermoWorks MicroThermo 2 och sond och registrerades med LabChartPro i realtid.
Poincar-mätningar och suck-och apnea-frekvens bestämdes med användning av 1 min rörelsefria spår från varje andningstillstånd. Suckar definierades som ett andetag med amplitud på minst dubbelt så stor som det genomsnittliga andetaget. Apneas definierades som ett interbreath interval (IBI) minst dubbelt så stort som det genomsnittliga IBI. Variationskoefficienten (CV) för IBI och amplitud beräknades också från samma 1-min spårkompilering av varje andningstillstånd (standardfel IBI eller amplitud/medel IBI eller amplitud).
statistik
pletysmografi
resultat (rr, VT, VE, VO2, VE/VO2, antal apneas och suckar och CVs av IBI och amplitud) för rumsluft och hyperkapnisk data jämfördes mellan DBH-Cre; RC::P_hM4D-kohorter och syskonkontroller med hjälp av en linjär regressionsmodell med blandade effekter med djurtyp (experimentell vs kontroll) och CNO-administrering (före vs efter injektion) som fasta effekter och djur-ID som en slumpmässig effekt. Temperaturdata jämfördes med användning av en linjär regressionsmodell med blandade effekter med djurtyp (experimentell kontra kontroll) som en fast effekt. En p < 0,05 användes för att indikera statistisk signifikans, och enskilda datapunkter, medelvärde och standardfel för medelvärdet visas på alla diagram.
vätskekromatografi-masspektrometri
det erhållna området under toppen för varje prov normaliserades av den interna kontrollen och sedan till vehikel innan statistisk analys utfördes. Oparat t-test användes för att jämföra det relativa överflödet av klozapin och CNO i varje vävnad efter koncentrationsgrupp.
Etikuttalande
alla experiment godkändes av Institutional Animal Care and Use Committee of Baylor College of Medicine. Experimenten överensstämde med nationella standarder för vård och användning av försöksdjur som fastställts av Association for Assessment and Accreditation of Laboratory Animal Care.
Författarbidrag
JS, FS-M, MC-M och RR utformade och utformade experimenten. JS, FS-M och PZ utförde experimenten och bidrog till att analysera data. JS, FS-M, VM, MC-M och RR skrev tidningen.
finansiering
denna studie stöddes av bidrag R01HL130249 och R01HL130249-S1 från National Heart, Lung and Blood Institute; Mars av Dimes Basil O ’ Connor Award; och McNair Medical Institute.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
vi tackar Dr Shaila K. Mani och Benjamin Arenkiel för hjälpsamma diskussioner. Vi tackar också Baylor College of Medicine Proteomics Core facility för utförandet av vätskekromatografi-masspektrometri.
Amiel, J., Dubreuil, V., Ramanantsoa, N., Fortin, G., Gallego, J., Brunet, J.-F., et al. (2009). PHOX2B i andningskontroll: lektioner från medfödd central hypoventilationssyndrom och dess musmodeller. Respir. Physiol. Neurobiol. 168, 125–132. doi: 10.1016 / j.resp.2009.03.005
CrossRef fulltext / Google Scholar
Andrews, N. Och File, S. E. (1993). Hantering av råttans historia modifierar beteendemässiga effekter av droger i det förhöjda Plus-labyrinttestet av ångest. EUR. J. Pharmacol. 235, 109–112. doi: 10.1016/0014-2999(93)90827-5
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S. och Roth, B. L. (2007). Utveckla låset för att passa nyckeln för att skapa en familj av G-proteinkopplade receptorer som aktiveras av en inert ligand. Proc. Natl. Acad. Sci. USA 104, 5163-5168. doi: 10.1073/pnas.0700293104
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ashby, C. R. och Wang, R. Y. (1996). Farmakologiska åtgärder av det atypiska antipsykotiska läkemedlet clozapin: en översyn. Synaps 24, 349-394. doi: 10.1002 / (SICI)1098-2396(199612)24:4<349::stöd-SYN5>3.0.CO; 2-D
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bahia, C. M. C. d. S. och Pereira, J. S. (2015). Obstruktiv sömnapnea och neurodegenerativa sjukdomar: en dubbelriktad relation. Dement. Neuropsychol. 9, 9–15. doi: 10.1590 / S1980-57642015DN91000003
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Biancardi, V., B Kubicego, K. C., Almeida, M. C. och Gargaglioni, L. H. (2008). Locus coeruleus noradrenerga neuroner och CO2 kör till andning. Pflugers Båge. – EUR. J. Physiol. 455, 1119–1128. doi: 10.1007 / s00424-007-0338-8
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Brust, Rd, Corcoran, ae, Richerson, GB, Nattie, E. och Dymecki, S. M. (2014). Funktionell och utvecklande identifiering av en molekylär subtyp av hjärnserotonerg neuron specialiserad för att reglera andningsdynamiken. Cell Rep. 9, 2152-2165. doi: 10.1016/j.celrep.2014.11.027
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Chang, W. H., Lin, S. K., Lane, H. Y., Wei, F. C., Hu, W. H., Lam, Y. W., et al. (1998). Reversibel metabolism av klozapin och klozapin N-oxid hos schizofrena patienter. Prog. Neuro-Psychopharmacol. Biol. Psykiatri 22, 723-739. doi: 10.1016 / S0278-5846(98)00035-9
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Chen, Y.-W., Das, M., Oyarzabal, E. A., Cheng, Q., Plummer, N. W., Smith, K. G., et al. (2018). Genetisk identifiering av en population av noradrenerga neuroner inblandade i dämpning av stressrelaterade svar. Mol. Psykiatri 1-16. doi: 10.1038 / s41380-018-0245-8
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kawasaki, H., Oya, H., Buzza, C., et al. (2015). Andning hämmas när anfall sprids till amygdala och vid Amygdala stimulering. J. Neurosci. 35, 10281–10289. doi: 10.1523/JNEUROSCI.0888-15. 2015
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2013). Rädsla och panik hos människor med bilateral amygdala skada. Nat. Neurovetenskap. 16, 270–272. doi: 10.1038 / nn.3323
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Fitton, A. och Heel, R. C. (1990). Klozapin. En översyn av dess farmakologiska egenskaper och terapeutisk användning vid schizofreni. Droger 40, 722-747. doi: 10.2165/00003495-199040050-00007
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2018). Tystnad av hypoglossala motoneuroner leder till sömnstörande andning i magra möss. Front. Neurol. 9:962. doi: 10.3389/fneur.2018.00962
CrossRef Fulltext / Google Scholar
Gargaglioni, L. H., Hartzler, L. K., och Putnam, R. W. (2010). Locus coeruleus och central kemosensitivitet. Respir. Physiol. Neurobiol. 173, 264–273. doi: 10.1016 / j.resp.2010.04.024
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Gomez, J. L., Bonaventura, J., Lesniak, W., Mathews, W. B., Sysa-Shah, P., Rodriguez, L. A., et al. (2017). Kemogenetik avslöjade: DREADD beläggning och aktivering via konverterad clozapin. Vetenskap 357, 503-507. doi: 10.1126/vetenskap.Aan2475
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Guettier, J. M. I., li, J. H., Rosemond, E., et al. (2009). En kemisk-genetisk metod för att studera g-proteinreglering av betacellfunktion in vivo. Proc. Natl. Acad. Sci. USA 106, 19197-19202. doi: 10.1073/pnas.0906593106
CrossRef fulltext / Google Scholar
Hakim, F., Gozal, D. och Kheirandish-Gozal, L. (2012). Sympatiska och katekolaminerga förändringar i sömnapnea med särskild tonvikt på barn. Front. Neurol. 3:7. doi: 10.3389/fneur.2012.00007
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Hennessy, M. L., Corcoran, A. E., Brust, R. D., Chang, Y., Nattie, E. E. och Dymecki, S. M. (2017). Aktivitet av Tachykinin1-Uttryckande Pet1 raphe-neuroner modulerar respiratorisk kemoreflex. J. Neurosci. 37, 1807–1819. doi: 10.1523/JNEUROSCI.2316-16. 2016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Huckstepp, R. T. R., Cardoza, K. P., Henderson, L. E. och Feldman, J. L. (2015). Roll parafacial kärnor i kontroll av andning hos vuxna råttor. J. Neurosci. 35, 1052–1067. doi: 10.1523/JNEUROSCI.2953-14. 2015
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ilg, A.-K., Enkel, T., Bartsch, D., och B Urachner, F. (2018). Beteendeeffekter av akut systemisk lågdos clozapin hos vildtypsråttor: konsekvenser för användningen av DREADDs i beteendemässig neurovetenskap. Front. Uppför dig. Neurovetenskap. 12:173. doi: 10.3389/fnbeh.2018.00173
CrossRef fulltext / Google Scholar
Isom, G. E. och Elshowihy, R. M. (1982). Interaktion av akut och kronisk stress med andning: modifiering av naloxon. Pharmacol. Biochem. Uppför dig. 16, 599–603. doi: 10.1016/0091-3057(82)90422-1
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Jann, MW, Lam, yw och Chang, Wh (1994). Snabb bildning av klozapin hos marsvin och människa efter administrering av klozapin-N-oxid. Båge. Int. Pharmacodyn. Där. 328, 243–250.
PubMed Abstrakt / Google Scholar
Joober, R. (2010). Clozapin: en distinkt, dåligt förstådd och underutnyttjad molekyl. J. Psykiatri Neurosci. 35, 147–149. doi: 10.1503/jpn.100055
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kinkead, R., Dupenloup, L., Valois, N. och Gulemetova, R. (2001). Stressinducerad dämpning av det hyperkapniska ventilationssvaret hos vakna råttor. J. Appl. Physiol. 90, 1729–1735. doi: 10.1152 / jappl.2001.90.5.1729
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Klein, D. F. (1993). Falska kvävningslarm, spontana panik och relaterade tillstånd. En integrerad hypotes. Båge. Gen. Psykiatri 50, 306-317. doi: 10.1001/archpsyc.1993.01820160076009
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Korsak, A., Sheikhbahaei, S., Machhada, A., Gourine, A. V. och Huckstepp, R. T. R. (2018). Parafaciala neurons roll i kontrollen av andning under träning. Sci. Rep. 8: 400. doi: 10.1038 / s41598-017-17412-z
CrossRef fulltext / Google Scholar
Lapin, I. P. (1995). Endast kontroller: effekt av hantering, skeninjektion och intraperitoneal injektion av saltlösning på beteende hos möss i en förhöjd Plus-labyrint. J. Pharmacol. Toxicol. Metoder 34, 73-77. doi: 10.1016 / 1056-8719 (95)00025-D
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Lavezzi, A. M., Alfonsi, G. och Matturri, L. (2013). Patofysiologi av det mänskliga locus coeruleus-komplexet i Foster / neonatal plötslig oförklarlig död. Neurol. Res. 35, 44-53. doi: 10.1179 / 1743132812Y. 0000000108
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2016). Clozapine N-oxid administrering ger beteendemässiga effekter i long-evans råttor: konsekvenser för att utforma DREADD experiment. eNeuro 3, 1-14. doi: 10.1523 / ENEURO.0219-16. 2016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Madisen, L., Zwingman, T. A., Sunkin, S. M., Oh, S. W., Zariwala, H. A., Gu, H., et al. (2010). En robust och hög genomströmning Cre rapportering och karakterisering system för hela musen hjärnan. Nat. Neurovetenskap. 13, 133–140. doi: 10.1038 / nn.2467
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Mahler, S. V. och Aston-Jones, G. (2018). CNO ont? Överväganden för användning av dreadds i beteende neurovetenskap. Neuropsykofarmakologi 43, 934-936. doi: 10.1038/npp.2017.299
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2018). DREADD-agonisten clozapin N-oxid (CNO) metaboliseras omvänd till klozapin och producerar klozapinliknande interoceptiva stimulanseffekter hos råttor och möss. Sci. Rep. 8, 3840. doi: 10.1038 / s41598-018-22116-z
CrossRef fulltext / Google Scholar
Misslin, R., Herzog, F., Koch, B. Och Ropartz, P. (1982). Effekter av isolering, hantering och nyhet på hypofysen-binjurssvar i musen. Psykoneuroendokrinologi 7, 217-221. doi: 10.1016/0306-4530(82)90015-4
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Nakamura, A., Zhang, W., Yanagisawa, M., Fukuda, Y. och Kuwaki, T. (2007). Vaksamhet tillståndsberoende dämpning av hyperkapnisk kemoreflex och överdriven sömnapnea i orexin knockout-möss. J. Appl. Physiol. 102, 241–248. doi: 10.1152 / japplphysiol.00679.2006
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Obonai, T., Yasuhara, M., Nakamura, T. och Takashima, S. (1998). Katekolaminneuroner förändras i hjärnstammen hos offer för plötsligt spädbarnsdöd. Pediatrik 101, 285-288. doi: 10.1542/peds.101.2.285
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ozawa, Y., Takashima, S. och Tada, H. (2003). Alfa2-adrenerga receptorsubtypförändringar i hjärnstammen i det plötsliga spädbarnsdödssyndromet. Tidig Hum. Dev. 75 (Suppl.), S129-S138. doi: 10.1016/j.pathophys.2004.01.015
CrossRef fulltext / Google Scholar
Padovan-Hernandez, Y., och Knackstedt, L. A. (2018). Dosberoende minskning av kokaininducerad rörelse genom clozapin-N-oxid hos råttor med en historia av kokain självadministrering. Neurovetenskap. Lett. 674, 132–135. doi: 10.1016/j.neulet.2018.03.045
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2011). Andning utan CO (2) kemosensitivitet i villkorliga Phox2b-mutanter. J. Neurosci. 31, 12880–12888. doi: 10.1523/JNEUROSCI.1721-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Raper, J., Morrison, R. D., Daniels, J. S., Howell, L., Bachevalier, J., Wichmann, T., et al. (2017). Metabolism and distribution of clozapine-N-oxide: implications for nonhuman primate chemogenetics. ACS Chem. Neurosci. 8, 1570–1576. doi: 10.1021/acschemneuro.7b00079
PubMed Abstract | CrossRef Full Text | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Soriano, L. P., Nattie, E. E., and Dymecki, S. M. (2013). Egr2-neuroner kontrollerar det vuxna respiratoriska svaret på hyperkapnia. Hjärnan Res. 1511, 115-125. doi: 10.1016/j.brainres.2012.12.017
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2011). Nedsatt andnings-och kroppstemperaturkontroll vid akut serotonerg neuroninhibering. Vetenskap 333, 637-642. doi: 10.1126/vetenskap.1205295
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Ryabinin, A. E., Wang, Y. M. och Finn, D. A. (1999). Olika nivåer av FOS-immunreaktivitet efter upprepad hantering och injektionsspänning i två inavlade stammar av möss. Pharmacol. Biochem. Uppför dig. 63, 143–151. doi: 10.1016 / S0091-3057(98)00239-1
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Snyder, B., Shell, B., Cunningham, jt och Cunningham, rl (2017). Kronisk intermittent hypoxi inducerar oxidativ stress och inflammation i hjärnregioner associerade med neurodegeneration i tidigt stadium. Physiol. Rep. 5, 1-13. doi: 10.14814 / phy2.13258
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Sun, JJ, Huang, T.-W., Neul, Jl och Ray, rs (2017). Embryonala bakhjärnmönstringsgener avgränsar distinkta hjärt-respiratoriska och metaboliska homeostatiska populationer hos vuxna. Sci. Rep. 7: 9117. doi: 10.1038 / s41598-017-08810-4
CrossRef fulltext / Google Scholar
Sun, JJ och Ray, rs (2017). Tg (Th-Cre)FI172Gsat (Th-Cre) definierar neuroner som krävs för fullständiga hyperkapniska och hypoxiska reflexer. Biol. Öppet 6, 1200-1208. doi: 10.1242 / bio.026823
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2014). Sängkärnan i stria terminalis är kritisk för ångestrelaterat beteende som framkallas av CO2 och acidos. J. Neurosci. 34, 10247–10255. doi: 10.1523/JNEUROSCI.1680-14. 2014
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Valentino, R. J. och Van Bockstaele, E. (2008). Konvergent reglering av locus coeruleus-aktivitet som ett adaptivt svar på stress. EUR. J. Pharmacol. 583, 194–203. doi: 10.1016/j.ejphar.2007.11.062
CrossRef fulltext / Google Scholar
Verbraecken, J. och McNicholas, W. T. (2013). Andningsmekanik och ventilationskontroll vid överlappningssyndrom och fetma hypoventilation. Respir. Upp 14: 132. doi: 10.1186/1465-9921-14-132
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Viemari, J.-C. (2008). Noradrenerg modulering av det respiratoriska neurala nätverket. Respir. Physiol. Neurobiol. 164, 123–130. doi: 10.1016 / j.resp.2008.06.016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Winter, A., Ahlbrand, R., Naik, D. och Sah, R. (2017). Differentiell beteendekänslighet för koldioxid (CO2) inandning hos råttor. Neurovetenskap 346, 423-433. doi: 10.1016/j.neurovetenskap.2017.01.003
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2009). Amygdala är en kemosensor som upptäcker koldioxid och acidos för att framkalla rädslabeteende. Cell 139, 1012–1021. doi: 10.1016/j.cell.2009.10.029
PubMed Abstract | CrossRef Full Text | Google Scholar