- introduktion
- material och metoder
- skörd av oocyter och cRNA-mikroinjektion
- isolering av Membranfraktioner och Immunoblotting
- immunohistokemi
- Frysfrakturelektronmikroskopi
- parad-Oocytanalys och kvantifiering av kontaktområden
- hydrostatisk Tryckimpulsanalys
- Kapratinkubation
- statistisk analys
- resultat
- uttryck av Claudin-5 och Integration i X. laevis Oocytplasmamembran
- fläckar av snäva Korsningssträngar är synliga i Claudin-5 som uttrycker oocyter
- parad Oocytanalys för analys av Claudin Trans-interaktion
- hydrostatisk Tryckimpulsanalys avslöjar Claudin-specifik korsning av Oocytpar
- inkubation med kaprat
- diskussion
- Claudins bidrar till starkare vidhäftningsegenskaper
- slutsats
- datatillgänglighet uttalande
- Etikuttalande
- Författarbidrag
- finansiering
- intressekonflikt
- erkännanden
- tilläggsmaterial
introduktion
den täta korsningsproteinfamiljen är avgörande för cellfysiologi eftersom brist eller försämring är förknippad med sjukdomar och dysfunktion hos många organ och vävnader, som visas t.ex. i innerörat (Wilcox et al., 2001; Florian et al., 2003), njure (Konrad et al., 2006; g Avsugningar et al., 2009), mag-tarmkanalen (Resnick et al., 2005; Amasheh et al., 2009), epidermis (Furuse et al., 2002; Tebbe et al., 2002) och hjärnkapillärer (Nitta et al., 2003; Wolburg et al., 2003). Claudiner representerar en transmembranproteinfamilj bestående av minst 27 medlemmar (Mineta et al., 2011). Förutom deras fyra transmembran helixdomäner innehåller de två extracellulära slingor (ECL1 och ECL2), en kort N-terminal och en C-terminal (Suzuki et al., 2014). Specifika claudin-uttrycksmönster bestämmer och återspeglar epitelens selektiva permeabilitet, och claudin-proteins förmåga att interagera i cis (inom samma membran) och i trans (mellan membranen i den närliggande cellen) möjliggör bildandet av barriärformning och porbildande täta korsningssträngar (Van Itallie och Anderson, 2006).
Claudin-5 uttrycks starkt i kapillärendotel och dominerar den täta korsningen (TJ) i blod-hjärnbarriären (BBB) eftersom uttrycket är >100 gånger högre jämfört med någon annan claudin (Ohtsuki et al., 2007). Dessutom uttrycks det i en mängd olika epitelvävnader inklusive lunga (Soini, 2011), exokrina vävnader (Comper et al., 2009), tarm (Garcia-Hernandez et al., 2017) och urinvägarna (Koda et al., 2011). Claudin-5 orsakar emellertid en starkare barriär i hjärnkapillärer än i andra vävnader (Reinhold och Rittner, 2017) och dess funktion försämras vid neurodegenerativa och neuroinflammatoriska störningar (Greene et al., 2019). Därför är claudin-5 avgörande för att upprätthålla BBB. Men BBB är inte bara skyddande, det begränsar också de terapeutiska alternativen eftersom läkemedel hindras för att genomsyra denna barriär.
Nitta et al. (2003) rapporterade att BBB är mer permeabel för molekyler av 800 Da i storlek i claudin-5-bristande möss jämfört med vildtypsmöss (Nitta et al., 2003). Detta var i enlighet med transfektionsexperiment som demonstrerade en tätningseffekt av claudin-5 i Caco-2-cellmonolager (Amasheh et al., 2005).
en annan viktig barriärbildande claudin är claudin-3, som har rapporterats selektivt försegla barriären mot passage av joner av antingen laddning och oladdade lösta ämnen (Milatz et al., 2010). Det uttrycks också i den endoteltäta korsningen av hjärnkapillärer och dess funktionella förlust observeras i faser av mikrovågsinflammation, glioblastom och choroid plexus hos patienter med multipel skleros (Engelhardt et al., 2001; Wolburg et al., 2003).
barriäregenskaper kan ändras dynamiskt, som t. ex., inkubation med natriumkaprat visades snabbt och reversibelt minska transepitelialt elektriskt motstånd i den mänskliga tarmcellinjen HT-29/B6 (Krug et al., 2013). Natriumkaprat öppnar tillfälligt claudin – 5 innehållande barriärer vid snäva korsningar av epitel-och endotelceller (Del Vecchio et al., 2012).
detta indikerar att claudin-5 är ett lovande mål för förbättring av läkemedelsleverans i BBB.
i denna studie syftade vi till att använda det heterologa uttryckssystemet för Xenopus laevis oocyter (Vitzthum et al., 2019) för analys av claudin-5 och claudin-3 interaktion och pertubation. På grund av bristen på endogena cell-cell-kontakter möjliggör detta enda celluttryckssystem analys av specifika claudiner utan störning av andra täta korsningsproteiner.
material och metoder
skörd av oocyter och cRNA-mikroinjektion
oocyter samlades in från vuxna kvinnliga afrikanska klogrodor genom kirurgisk laparotomi. För anasthesia, 0.2% MS222 (etyl-3-aminobensoatmetansulfonat, Sigma-Aldrich, Taufkirchen, Tyskland) användes som en badlösning i 5-10 min vid 20 kcal C. När kirurgisk anasthesi uppnåddes gjordes hud-och bukmuskelinsnitt och äggstocksmassan exterioriserades och ovariell vävnad avlägsnades. Isoleringen av oocyter utfördes genom enzymatisk digestion vid rumstemperatur i 90 minuter i 1,5 mg/ml kollagenas Fisher Bioreagenter BP2649-1 (Fisher Scientific, Schwerte, Tyskland) upplöst i oocyt Ringer solution (ORi) såsom beskrivits av Vitzthum et al. (2019). Follikulära celler avlägsnades genom inkubation i Ca2 + – fri ORi innehållande (i mM): NaCl (90), KCl (1), EGTA (trietylenglykoldiamintetraättiksyra) (1), 5 HEPES (5); pH 7,4 för 10 min på en mekanisk skakare med 50 rpm. Oocyter i steg V och VI (>1000 occurm) injicerades (Nanoliter 2010, World Precision Instruments, Sarasota, FL, USA) med 1 ng cRNA som kodar för humant claudin-5, claudin-3 eller RNase-fritt vatten som kontroller. Injektionsvolymen var 50,6 nl per oocyt. Efter injektion inkuberades oocyter vid 16 kcal C i ORi 3 dagar för proteinuttryck.
isolering av Membranfraktioner och Immunoblotting
tio injicerade oocyter poolades för western blot-analys och resuspenderades i 500 occl homogeniseringsbuffert innehållande (i mM) MgCl2 (5), NaH2PO4 (5), EDTA (etylendiamintetraättiksyra) (1), sackaros (80) och Tris (Tris(hydroximetyl)aminometan) (20); pH 7.4. Oocytextrakt centrifugerades två gånger vid 200 rpm för 10 min vid 4 CCB för att kassera cellskräp. Supernatanten centrifugerades vid 13 000 rpm för 30 min vid 4 CCB för att pelletera cellmembranet såsom beskrivits av Leduc-Nadeau et al. (2007). Pellets resuspenderades i 80 occl homogeniseringsbuffert. Proteinkvantifiering gjordes kolorimetriskt med Pierce 600 nm Proteinanalyssats (Thermo Fisher Scientific, Hennigsdorf, Tyskland) enligt tillverkarens instruktion i en 96 brunnsplatta. Plattläsaren (PerkinElmer enspire multimode Plate Reader, Waltham, MA, USA) justerades till 562 nm och bovint serumalbumin Standard (Thermo Fisher Scientific, Hennigsdorf, Tyskland) som sträcker sig från 125 till 2000 kg/ml användes för utvärdering. Före immunoblotting blandades proverna med 4 Aci laemmli-buffert (Bio-Rad Laboratories, Munich, Tyskland), laddade på en 10% SDS-polyakrylamidgel och elektroforerade. För proteinöverföring användes PVDF-membran och blockerades i 5% torrmjölk utan fett i Trisbuffrad saltlösning i 120 minuter. Proteiner detekterades genom immunoblotting med användning av primära antikroppar upphöjda mot claudin – 3 eller claudin-5 (invitrogen #35-2500, #34-1700, #34-1600, Life Technologies, Carlsbad, CA, USA).
peroxidas-konjugerade get-anti-kanin-och anti-musantikroppar (#7074, #7076 Cellsignaleringsteknik, Danvers, MA, USA) användes för att binda till de primära antikropparna och inkuberades därför i minst 45 minuter vid rumstemperatur. För detektion användes Clarity Western ECL Blotting Substrate (#1705061, Bio-Rad Laboratories GmbH, Munich, Tyskland) och signaler visualiserades av ett ChemiDoc MP-system (Bio-Rad Laboratories).
immunohistokemi
injicerade oocyter fixerades i 4% PFA (16% paraformaldehyd, E15700, Science Service, Munich, Tyskland) för 4 h vid rumstemperatur följt av dehydreringsgradient från 70% etanol till xylol (Carl Roth, Karlsruhe, Tyskland) inom 48 h. prover inbäddades i paraffin och tvärsnittades (5 kg) med användning av en Leica RM 2245 mikrotom (Leica Microsystems Heidelberg, Tyskland). Strax före immunhistokemisk behandling avlägsnades paraffin via xylol till etanolgradient. Icke-specifika bindningsställen blockerades med 5% getserum i fosfatbuffrad saltlösning och inkuberades med samma primära antikroppar som för immunoblotting. Prover inkuberades med sekundära antikroppar Alexa Fluor – 488 get anti-kanin och Alexa Fluor-594 get anti-mus (Life Technologies, Carlsbad, CA, USA) och undersöktes genom konfokal laserskanning immunofluorescensmikroskopi (LSM 710, Zeiss, Oberkochen, Tyskland).
Frysfrakturelektronmikroskopi
Frysfrakturelektronmikroskopi utfördes som rapporterats nyligen (Greene et al., 2019). För fixering inkuberades injicerade oocyter i glutaraldehyd (2,5% i 0,1 M cacodylatbuffert) över natten vid 4 msk C. Efter tvättning med cacodylatbuffert bereddes oocyter för frysfrakturering. Proverna kryoskyddades i 30% glycerol och frystes i flytande kväve. Efter sprickbildning och skuggning med platina och kol (BAF400D; Balzers, Liechtenstein) avlägsnades återstående organiskt material genom en natriumhypoklorit tvätt. Oocyter analyserades i ett transmissionselektronmikroskop (EM-10, Zeiss, Oberkochen, Tyskland) och fotograferades med en digitalkamera (tr Kazakndle GmbH). Morfometrisk analys av de snäva korsningssträngarna utfördes vid en förstoring av 20 000 kg.
parad-Oocytanalys och kvantifiering av kontaktområden
Mannitol implementerades för att krympa de injicerade oocyterna och möjliggöra ett mekaniskt avlägsnande av vitellinmembranet med pincett utan att skada plasmamembranet. 5-10 oocyter placerades i en petriskål (35 mm diameter, Thermo Fisher, Henningsdorf, Tyskland, #153066) fylld med ORi. Mannitol tillsattes och löstes tills hypertonisk krympning av cellerna uppnåddes (cirka 400 mOsmol/l under 10 min). Efter manuell devitellinisation överfördes oocyter omedelbart till en 24 brunnsplatta (1. 86 cm2 yta, TPP Techno plastprodukter, Trasadingen, Schweiz, # 92024) innehållande 2 ml ORi. I varje brunn grupperades två celler försiktigt genom att trycka dem ihop med en Pasteurpipett (1 ml, Thermo Fisher, Henningsdorf, Tyskland, #PP88SB) och en bulbous sond.
Oocytpar av claudin-5-Uttryckande (cldn5 − cldn5), claudin-3-Uttryckande (cldn3 − cldn3), claudin-3 och claudin-5 coexpressing (cldn3,5 − cldn3,5) och kontrollocyter (kontroll − kontroll) hölls tillsammans i upp till 48 h I ORi vid 16 kcal C.
Ljusfältmikroskopi användes för kvantifiering av kontaktområdet för klustrade oocyter efter 1, 24 och 48 h. bilder av Na-oocytpar i 24 brunnskulturrätter togs vid dessa tidpunkter med hjälp av ett Leica dmi6000 B-mikroskop (Leica Microsystems, Wetzlar, Tyskland). Diametern på kontaktytan mättes med användning av mikron skala (LAS-AF 3.2.0). Kontaktytor anses vara cirkulära och därmed beräknades kontaktytan med hjälp av cirkelekvationen A=R2.
hydrostatisk Tryckimpulsanalys
vitellinmembran avlägsnades mekaniskt som beskrivits tidigare och oocyter grupperades analagt till den parade oocytanalysen. Dessutom testades blandade oocytpar (kontroll–cldn5 och kontroll–cldn3) i hydrostatisk tryckimpuls (HPI) – analys. Efter 24 timmars stabilisering skapades en definierad hydrostatisk impuls med en enda kanal elektronisk pipett (EE-300R, Eppendorf Research Pro, programvaruversion 2.06.00, Hamburg, Tyskland).
oocyter förvarades i 24 brunnsplattor innehållande 2 ml ORi och central positionering kontrollerades före applicering av pipetteringsvolymen 250 occl ORi. Doseringshastigheten var enhetlig inställd på maximal hastighet, vilket motsvarade en doseringshastighet på 0,9 s. dessutom applicerades vinkeln (45 kg) och appliceringsavståndet (1,3 cm) enhetligt. Omgivningstryck, viskositet hos ORi och diameter av pipettspetsöppningen hölls under konstanta förhållanden. Ljusfältmikroskopi användes för kvantifiering av kontaktområden 30 min efter det hydrostatiska trycket applicerades och jämfördes med kontaktområden före applicering. Den experimentella inställningen beskrivs i Figur 1.
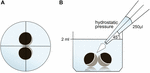
Figur 1. Experimentell inställning av hydrostatisk tryckimpulsanalys (a) schematisk toppvy av brunnen: Central positionering av oocytparet kontrollerades före applicering av det hydrostatiska trycket. (B) schematisk sidovy av brunnen: 250 occl ORi tillsattes med användning av en enda kanal elektronisk pipett. Doseringshastigheten var enhetlig inställd på maximal hastighet. Vinkeln (45 kcal) och appliceringsavståndet applicerades enhetligt. Omgivningstryck, viskositet av ORi och diameter av pipettspetsöppningen hölls under konstanta betingelser.
Kapratinkubation
för kapratinkubation tillsattes natriumkaprat (#C4151, Sigma Aldrich, Taufkirchen, Tyskland) i Slutliga koncentrationer på 50, 100 och 500 kg, eller ORi som referensgrupp, till oocyterna 24 timmar efter parning. Oocyter hölls i 24 brunnsplattor innehållande 2 ml ORi och kapratlösning löstes i en definierad tillsatsvolym av 250 oc per brunn. Bredden på kontaktytan kvantifierades vid 30, 60 och 120 min efter tillsats.
statistisk analys
statistisk analys utfördes med JMP Pro 14.0.0 (NC, USA). Data presenteras som medianer och visas som procentuell förändring baserat på den grupperade kombinationen vid de första undersökningspunkterna. Figurerna 4, 5 presenteras som lådor, som visar den första kvartilen (25 procent), medianen (50 procent) och den andra kvartilen (75 procent). Morrhåren dras ner till 10: e percentilen och upp till 90: e percentilen. Normalfördelningen kontrollerades med hjälp av Shapiro–Wilk-test.
Kruskal–Wallis-test användes för multipel jämförelse, följt av en Dunn-Bonferroni-korrigering. p-värden anges som kontinuerliga tal.
resultat
uttryck av Claudin-5 och Integration i X. laevis Oocytplasmamembran
för att testa det framgångsrika uttrycket och integrationen av det täta korsningsproteinet claudin-5 i oocytplasmamembranet, 3 dagar efter injektion av claudin-5 cRNA, analyserades membranfraktioner genom immunoblotting. Alla prover från tre enskilda djur (d1–d3) avslöjade claudin-5-specifika signaler vid 23 kDa, medan RNAs-fria vatteninsprutade oocyter inte visade någon specifik signal för claudin-5-uttryck (figur 2a).
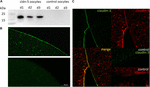
Figur 2. Detektion av heterologt uttryckta claudiner i Xenopus laevis oocyter. (A) Immunoblot-analys av tätt korsningsprotein claudin–5 i X. laevis-oocyter av tre djur (d1-d3). Cellmembranlysater utsattes för SDS-sida följt av immunoblot på PVDF-membran. Membranen inkuberades med primära antikroppar och sekundära peroxidas-konjugerade antikroppar (n = 3). (B) immunofluorescerande färgning avslöjade specifika claudin-5-signaler (grön) i oocytmembran av alla cRNA-injicerade oocyter, medan i vatteninjicerade kontroller detekterades inga claudin-specifika signaler i konfokalmikroskopi. Representativa bilder av oocyter härledda från tre djur. Skala bars: 50 oc. m. (C) immunofluorescerande färgning av claudin-5 och claudin-3-Uttryckande oocyter avslöjade specifika claudin-3-signaler (grön) och claudin-5-signaler (röd) i oocytmembran av cRNA-injicerade oocyter, medan i vatteninjicerade kontroller detekterades inga claudin-specifika signaler. Kolokalisering av uttryckta claudin-proteiner i oocytplasmamembranet avslöjas genom dubbel immunofluorescerande färgning (gul). Skala bar: 10 oc. m.
för visualisering av de uttryckta proteinerna i plasmamembranet utfördes immunhistokemiska fläckar och analyserades med konfokal laserskanningsmikroskopi (Figur 2B). Specifika signaler detekterades och fördelades jämnt över plasmamembranet hos claudin-5 som uttryckte oocyter. I enlighet med immunblot detekterades inga specifika signaler i kontrollocytplasmemembran.
således, efter injektion av cRNA, uttrycktes claudin-5 framgångsrikt och integrerades i plasmamembranet av X. laevis-oocyter.
Co-expression av claudin-3 och claudin-5 i oocytpar avslöjade specifika signaler för claudin-5 (röd) och claudin-3 (Grön) i båda cellerna (figur 2C). Oocytplasmemembran visade en fusion av de närliggande cellerna som tillhandahålls av direkt cldn3,5–cldn3,5-interaktion (gul).
oocytplasmamembran analyserades och visualisering av snäva korsningssträngar lyckades (Figur 3). Frysfrakturelektronmikroskopi visade fläckar av strängmorfologi i plasmamembranen hos claudin-5 injicerade oocyter, och strängorganisation av claudin-5-Uttryckande oocyter var mycket organiserad och av vinkelform (figur 3A). Täta korsningssträngar detekterades primärt i membranets protoplasmiska (P-) yta. Claudin – 3 injicerade oocyter visade rundade mycket organiserade snäva korsningssträngar som rapporterats tidigare (Vitzthum et al., 2019; figur 3B). Frysfrakturelektronmikroskopi av claudin – 3 och claudin-5 coexpressing oocyter avslöjade fibriller som båda bär egenskaper hos claudin-3 och claudin-5. Fibril strandarkitektur av coexpressing oocyter verkade både rundade och komplexa som claudin-3-Uttryckande oocyter, men också diskontinösa och mer vinklade som visat för claudin-5-Uttryckande celler (figur 3C). Kontrollocyter hade en typisk slät yta (figur 3D).
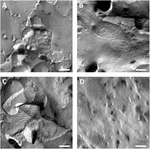
Figur 3. Frys fraktur elektronmikroskopi. (A) Frysfrakturelektronmikroskopi avslöjar tätt korsningsprotein cldn-5 som ett nät av vinklade diskontinösa fibriller i rader i Xenopus laevis-oocyter. (B) Frysfrakturelektronmikroskopi avslöjar tätt korsningsprotein cldn-3 som ett nät av rundade fibriller i X. laevis-oocyter. (C) Frysfrakturelektronmikroskopi av claudin-3 och claudin-5 samuttryckande oocyter avslöjar fibriller som båda har egenskaper hos claudin-3 och claudin-5. (D) vatteninsprutade kontrollocyter har en slät yta. Representativa bilder av oocyter härledda från tre djur. Skala bar: 250 nm.
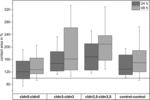
Figur 4. Mediankontaktområden för klustrade oocytkombinationer cldn5-cldn5, cldn3–cldn3,cldn3,5–cldn3, 5 och kontrollkontroll 24 och 48 timmar efter kluster i % av de initiala kontaktområdena kort efter kluster (n = 8-38).

Figur 5. Kontaktområden av claudin-5, claudin-3 och coexpressing claudin-3 och claudin-5 oocyter i hydrostatisk tryckimpuls (HPI) utmaning efter stabiliseringsperiod och 30 min efter HPI (n = 16-70, *p < 0,05, **p < 0.01, Kruskal-Wallis följt av en Dunn-Bonferroni korrigering).
parad Oocytanalys för analys av Claudin Trans-interaktion
alla grupperade kombinationer visade en tidsberoende ökning av kontaktytan under den uppmätta tidsperioden (Figur 4 och Tabell 1).
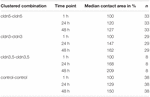
Tabell 1. Oocytkontaktområden inom 48 timmar efter kluster.
kontaktområdet för vatteninsprutade kontrollocyter ökade till 129% efter 24 timmar och 150% efter 48 timmar. Klustrade par oocyter som uttryckte claudin – 3 visade också en ökning av kontaktområdena till 147% (24 h) och 162% (48 h). Grupperade par av oocyter som samuttrycker claudin – 3 och claudin-5 visade kontaktområden på 168% (24 h) och 209% (48 h). Grupperade par oocyter som uttryckte claudin-5 ensam visade kontaktområden på 120% (24 h) och 127% (48 h). Därför var kontaktområdena i alla testade kombinationer jämförbara.
hydrostatisk Tryckimpulsanalys avslöjar Claudin-specifik korsning av Oocytpar
i ett separat tillvägagångssätt, oocyter som uttrycker claudin-3 eller claudin-5 eller samuttryckande båda claudinerna grupperades efter mekanisk devitellinisering. Oocyter utmanades genom att använda en HPI och kontaktområden mättes och beräknades 30 min efter utmaning och jämfördes med initiala områden efter 24 h stabiliseringsperioden (Figur 5 och Tabell 2). Efter hydrostatisk tryckutmaning minskade kontaktområdet för vatteninsprutade kontrollocyter till 89%. Klustrade par oocyter som uttrycker claudin-5, claudin-3 eller samuttryckande claudin-3 och claudin-5 behöll större kontaktområden (97%, p = 0,0235; 96%, p = 0,003; 98%, p = 0,0253). Kontaktområdena för blandade vatteninsprutade kontrollocyter och claudin-Uttryckande oocyter (kontroll–cldn5 och kontroll–cldn3) skilde sig inte signifikant från kontrollocyter (93%, p = 0,2900; 83%, p = 0,4455).
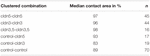
Tabell 2. Hydrostatisk tryckimpulsanalys.
inkubation med kaprat
i ett pilotinkubationsexperiment injicerades oocyter och parades i kombinationer som antingen uttryckte claudin-5 (cldn5–cldn5) eller injicerades med RNAs–fritt vatten som kontroller (kontroll-kontroll). Par inkuberades med slutliga natriumkapratkoncentrationer på 50, 100 eller 500 occurm. Inkubationen av oocytpar med ORi fungerade som en referensgrupp. Oocyter grupperades och efter 24 timmars stabilisering började inkubation och kontaktbredder mättes 30, 60 och 120 min efter tillsats (kompletterande figur S1).
tillsatsen av ORi resulterade i en initial minskning av kontaktområdena både i claudin-Uttryckande och vatteninsprutade oocytpar som dispergeras 60 eller 120 min efter tillsats. Detta skisseras av den paraboliska formen av anslutningslinjen mellan mediankontaktområdena över tiden (röda kurvor i Tilläggsfigur S1). Men, inkubation med 100 och 500 µM natrium caprate ökade kontaktytor något (100 µM) eller starkt (500 µM) 30 min efter caprate förutom från 5,1 × 105 5,2 × 105 µm2 och 4,3 × 105 4,9 × 105 µm2.
diskussion
i den aktuella studien använde vi den klassiska modellen för transportörer och mänsklig sjukdomsmodellering (Tammaro et al., 2009; Nenni et al., 2019), X. laevis-oocyterna, för en djupgående analys av claudin-5-interaktion och funktionellt bidrag till korsningsförseglingen. För detta ändamål etablerades ett nytt tillvägagångssätt, införande av en HPI för utmanande interaktion inom kontaktområdet för grupperade Xenopus-oocyter.
Claudins bidrar till starkare vidhäftningsegenskaper
i enlighet med tidigare resultat från Vitzthum et al. (2019) ledde enstaka claudiner uttryckta i oocyter inte till en ökning av interaktionskontaktområden jämfört med kontrollocyter. Immunoblot och immunohistokemisk visualisering visade emellertid det framgångsrika uttrycket och integrationen i X. laevis oocytplasmamembran. Vidare möjliggjorde användningen av konfokal laserskanningsmikroskopi en exakt lokalisering av de uttryckta claudinerna i plasmamembranet när pinhålet blockerade fluorescens utanför fokus. En kvantifiering av immunhistokemiska signaler eftersträvades inte, eftersom antikropparnas affinitet för bindning av deras mål skiljer sig åt.
Vedula et al. (2009) använde en mikropipett aspirationsteknik för att undersöka aspekter av claudin-claudin-interaktion med L-fibroblaster transfekterade med GFP-taggade occludin, cldn-1 och cldn-2. Separationskraften som behövdes för att lossa två celler från varandra var större i cldn-1 och cldn-2 transfekterade celler (2,8 respektive 2,3 nn). Även om detta tillvägagångssätt kan verka lovande för claudin-claudin-interaktionsanalys av att uttrycka oocyter, preliminära tester visade att en avskiljning av klustrade oocyter inte är möjlig utan störning av oocytplasmemembranen.
därför, som ett nytt tillvägagångssätt, mättes anslutningarnas kraft med en HPI. Även om HPI inte ger en kvantifiering av separationskraften i absoluta värden (t. ex. i Newtons), tillåter den en snabb och kostnadseffektiv analys av claudin-interaktionen utan störning av andra täta korsningsproteiner (t. ex., occludin, tricellulin, sylt-A). Claudiner bidrar till korsningen av oocytpar eftersom de visar ett större kontaktområde jämfört med vatteninsprutade oocyter efter HPI. Detta indikerar stark homofil trans-interaktion mellan de claudin-Uttryckande cellerna.
Strand fibril arkitektur är specifik för enstaka claudins (Colegio et al., 2002). I frysfrakturelektronmikroskopi rapporterades claudin-3 att montera ett mer rundat strängnät i slingformer i Xenopus-oocyter (Vitzthum et al., 2019; Figur 3B) medan claudin-5 i vår studie bildade ett nätverk mer vinklat och ordnat i rader. Bilderna avslöjade att det täta korsningsproteinet claudin-5 bildar ett nät av fibriller i diskontinösa, vinkelformade rader i X. laevis-oocyter. Detta är i enlighet med claudin-5-strängar som är kända för att förekomma som kedjor av partiklar associerade med P-fasen (Piontek et al., 2011). I våra experiment verkade fibrillernas geometriska form inte ha någon effekt på oocyternas vidhäftningsegenskaper. I enlighet med detta rapporterades paracellulär resistens vara orelaterad med fibrilnummer och fibrilbildande egenskaper (Colegio et al., 2002). Dessutom har claudin – 3 och claudin-5 visat sig ha en liknande förmåga för homofil trans-interaktion i HEK293-celler (Piontek et al., 2011).
denna modell av X. laevis-oocyter, som uttrycker enstaka täta korsningsproteiner, möjliggör en observation av effekten av ämnen som natriumkaprat på bildandet av kontaktområden mellan klustrade oocyter. Det kan därför vara ett användbart verktyg för en tids-och kostnadseffektiv screening för ämnen som påverkar den täta korsningsbarriären.
natriumkapratkoncentrationer på 100 och 500 oc kan ge en skyddande effekt på claudin-5-Uttryckande oocyter, vilket resulterar i ökande kontaktområden efter 30 min inkubation.
Krug et al. (2013) visade att inkubation med natriumkaprat ledde till en snabb och reversibel minskning av transepitelial resistens i human tarmcellinje HT-29/B6. Vidare avslöjade konfokal laserskanningsmikroskopi en markant reduktion av claudin-5 i ht-29/B6-celler behandlade med medelkedjiga fettsyralauratet (Dittmann et al., 2014). Den första extracellulära slingan av claudiner (ECL1) är viktig för barriäregenskaperna hos den täta korsningen, medan den andra extracellulära slingan (ECL2) är involverad i strängbildning av trans-interaktion (Piontek et al., 2008; Rossa et al., 2014; Greene et al., 2019). Claudin-5-bindemedel riktade mot ECL1 eller ECL2 av claudin-5 kan inducera intracellulärt upptag av det täta korsningsproteinet och därigenom motverka claudin-5-trans-interaktioner i den täta korsningsförseglingen mellan intilliggande celler och lossa det paracellulära utrymmet (Hashimoto et al., 2017).
således antog vi att inkubation med natriumkaprat skulle leda till en minskning av kontaktområdet för klustrade claudin-5 som uttrycker X. laevis-oocyter. Oväntat ledde ökande koncentrationer av natriumkaprat (100 och 500 kg) till ökande kontaktområden av klustrade oocyter som uttryckte claudin-5 vilket indikerar en skyddande effekt av natriumkaprat på den täta korsningsförseglingen. Vidare beskrivs natriumkaprat för att inducera sammandragning av aktomyosin perijunktionell ring, vilket utvidgar det paracellulära utrymmet (Lindmark et al., 1998; Maher et al., 2009). Denna effekt är baserad på fosforylering av den regulatoriska lätta kedjan av myosin via en fosfolipas C-aktivering som leder till en klyvning av fosfatidylinositol 4,5-bisfosfat (PIP2) till inositoltrifosfat (IP3) och diacylglycerol (Tomita et al., 1995). Det snäva korsningskomplexet som är kopplat av ställningsproteinet ZO-1 till aktincytoskelettet omfördelas sedan från den täta korsningen till cytoplasman (Lindmark et al., 1998; Turner, 2000). Cytoarkitekturen hos X. laevis-oocyten är avgörande för cytoplasmatisk regionalisering under oogenes (Wylie et al., 1985). Fastän, före befruktning, tjp-1–genuttryck, uttrycks genen som kodar för ZO-1 endast 1,9 TPM i oocytstadier V-VI (Session et al., 2016). En minskad interaktion mellan claudinerna och cytoskeletala ställningen kan därför förklara det oväntade resultatet av kapratinkubationen. Är omfattningen av framtida studier för att verifiera den mekanistiska grunden för upptäckten.
en varianteffekt av natriumkaprat på den paracellulära permeabiliteten beskrevs emellertid tidigare i litteraturen. I Peyers Patchvävnad som tagits från tarmen hos vuxna grisar upptäcktes en liknande förstärkningseffekt. Claudin-5 ökades signifikant efter inkubation med 5 mM kaprat. I denna studie ledde kaprat till en signifikant högre transepitelial elektrisk resistans (TEER) i follikelassocierat epitel (Radloff et al., 2019).
slutsats
Sammanfattningsvis heterologt uttryck av det täta korsningsproteinet claudin-5 i X. laevis oocyter möjliggör nya insikter i enskilda claudins bidrag till cell-cellinteraktion och vidhäftningsegenskaper hos intilliggande celler. Således möjliggör användning av X. laevis tight junction-modellen för claudin-5 analys av BBB-komponenter i en encellig modell.
datatillgänglighet uttalande
de dataset som genereras för denna studie är tillgängliga på begäran till motsvarande författare.
Etikuttalande
djurbehandlingarna var i enlighet med riktlinjerna i tysk lagstiftning, med godkännande av djurskyddsansvarig för Freie Universit Jacobt Berlin och under ledning av Berlins Veterinary Health Inspectorate (Landesamt f jacobr Gesundheit und Soziales Berlin, tillstånd G0025/16).
Författarbidrag
alla författare har läst och godkänt manuskriptet. NB och SA designade, planerade och övervakade experimenten och skrev manuskriptet. NB, LS, VC, RK och PF-B utförde experimenten och dataanalysen.
finansiering
denna studie finansierades av Deutsche Forschungsgemeinschaft, bidrag nr. AM141 / 11-1 och H. Wilhelm Schaumann Stiftung.
intressekonflikt
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
erkännanden
vi tackar Martin Grunau, Gisela Manz, Katharina s Excepllig och Susanne Trappe för utmärkt teknisk hjälp. Vi erkänner stöd från Open Access Publication Initiative från Freie universitet i Berlin.
tilläggsmaterial
Tilläggsmaterialet för denna artikel finns online på: https://www.frontiersin.org/articles/10.3389/fphys.2020.00857/full#supplementary-material
figur S1 / kontaktområden av claudin-5 som uttrycker oocytpar i tuborim2 under inkubation med natriumkaprat i olika koncentrationer (n = 6-8 respektive) och vatteninjicerade oocyter som kontroller (n = 5-7 respektive).
Amasheh, S., Dullat, S., Fromm, M., Schulzke, jd, Buhr, Hj och Kroesen, aj (2009). Inflammerad påse slemhinna har förändrade snäva korsningar som indikerar återkommande inflammatorisk tarmsjukdom. Int. J. Colorect. Dis. 24, 1149–1156. doi: 10.1007 / s00384-009-0737-8
PubMed Abstrakt / CrossRef fulltext / Google Scholar
amasheh, S., Schmidt, T., Mahn, M., Florian, P., Mankertz, J., Tavalali, S., et al. (2005). Bidrag av claudin-5 till barriäregenskaper i täta korsningar av epitelceller. Cellvävnad Res. 321, 89-96. doi: 10.1007 / s00441-005-1101-0
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Colegio, O. R., Van Itallie, C., Rahner, C. och Anderson, J. M. (2002). Rollen av claudin extracellulära domäner i tight junction fibril arkitektur och paracellulär laddningsselektivitet. Mol Biol Cell. 13:286.
Google Scholar
C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C., C. (2009). Expressionsmönster av claudiner 5 och 7 skiljer fast pseudopapillär från pankreatoblastom, acinarcell och endokrina tumörer i bukspottkörteln. Är. J. Surg. Pathol. 33, 768–774. doi: 10.1097 / pas.0b013e3181957bc4
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2012). Natriumkaprat öppnar tillfälligt claudin-5-innehållande barriärer vid snäva korsningar av epitel-och endotelceller. Mol. Läkemedelsföretag. 9, 2523–2533. doi: 10.1021 / mp3001414
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Dittmann, I., Amasheh, M., Krug, S. M., Markov, A. G., Fromm, M. och Amasheh, S. (2014). Laurate genomsyrar den paracellulära vägen för små molekyler i tarmepitelcellsmodellen HT-29/B6 Genom att öppna de täta korsningarna genom reversibel omlokalisering av claudin-5. Läkemedelsföretag. Res Dordr. 31, 2539–2548. doi: 10.1007 / s11095-014-1350-2
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Engelhardt, B., Wolburg-Buchholz, K. och Wolburg, H. (2001). Inblandning av choroid plexus i inflammation i centrala nervsystemet. Microsc. Res.Techniq. 52, 112–129. doi: 10.1002/1097-0029(20010101)52:1<112::stöd-jemt13>3.0.co;2-5
CrossRef fulltext / Google Scholar
L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L., L. (2003). Claudins i de snäva korsningarna av stria vascularis marginella celler. Biochem. Biophys. Res.Co. 304, 5–10. doi: 10.1016 / s0006-291x(03)00498-4
CrossRef fulltext / Google Scholar
Furuse, M., Hata, M., Furuse, K. Han är en av de mest kända i världen. (2002). Claudin-baserade täta korsningar är avgörande för däggdjurens epidermala barriär: en lektion från claudin-1-bristfälliga möss. J. Cell Biol. 156, 1099–1111. doi: 10.1083 / jcb.200110122
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Garcia-Hernandez, V., Quiros, M. och Nusrat, A. (2017). Intestinala epitelklaudiner: uttryck och reglering vid homeostas och inflammation. Ann. N. Y. Acad. Sci. 1397, 66–79. doi: 10.1111/nyas.13360
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Greene, C., Hanley, N. och Campbell, M. (2019). Claudin-5: gatekeeper av neurologisk funktion. Vätskor Barr. CNS 16: 4321.
Google Scholar
G. A., D., Amasheh, S., Pfaffenbach, S., Richter, J. F., Kausalya, P. J., Hunziker, W., et al. (2009). Claudin – 16 påverkar transcellulär Cl – sekretion i MDCK-celler. J. Physiol. Lond. 587, 3777–3793. doi: 10.1113 / jphysiol.2009.173401
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Hashimoto, Y., Shirakura, K., Okada, Y., Takeda, H., Endo, K., Tamura, M., et al. (2017). Claudin-5-bindemedel förbättrar permeationen av lösta ämnen över blod-hjärnbarriären i en däggdjursmodell. J. Pharmacol. Exp. Där. 363, 275–283. doi: 10.1124/jpet.117.243014
PubMed Abstrakt / CrossRef fulltext / Google Scholar
koda, R., Zhao, L. N., Yaoita, E., Yoshida, Y., Tsukita, S., Tamura, A., et al. (2011). Nytt uttryck av claudin-5 i glomerulära podocyter. Cellvävnad Res. 343, 637-648. doi: 10.1007 / s00441-010-1117-y
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2006). Mutationer i den snäva korsningsgenen claudin 19 (CLDN19) är associerade med renalt magnesiumavfall, njursvikt och allvarligt okulärt engagemang. Är. J. Hum. Genet. 79, 949–957. doi: 10.1086/508617
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Krug, S. M., Amasheh, M., Dittmann, I., Christoffel, I., Fromm, M. och Amasheh, S. (2013). Natriumkaprat som en förstärkare av makromolekylpermeation över tricellulära täta korsningar av tarmceller. Biomaterial 34, 275-282. doi: 10.1016 / j.biomaterial.2012.09.051
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Leduc-Nadeau, A., Lahjouji, K., Bissonnette, P., Lapointe, J. Y. och Bichet, D. G. (2007). Utarbetande av en ny teknik för rening av plasmamembran från Xenopus laevis oocyter. Är. J. Physiol. Cell. 292, C1132-C1136.
Google Scholar
Lindmark, T., Kimura, Y. och Artursson, P. (1998). Absorptionsförbättring genom intracellulär reglering av tät korsningspermeabilitet med medelkedjiga fettsyror i Caco-2-celler. J. Pharmacol. Exp. Där. 284, 362–369.
Google Scholar
Maher, S., Leonard, T. W., Jacobsen, J. och Brayden, D. J. (2009). Säkerhet och effekt av natriumkaprat för att främja oral läkemedelsabsorption: från in vitro till kliniken. Adv. Läkemedel Leverera. Rev. 61, 1427-1449. doi: 10.1016 / j. addr.2009.09.006
PubMed Abstrakt / CrossRef fulltext / Google Scholar
milatz, S., Krug, S. M., Rosenthal, R., Gunzel, D., Muller, D., Schulzke, J. D., et al. (2010). Claudin – 3 fungerar som en tätningskomponent i den täta korsningen för joner av antingen laddning och oladdade lösta ämnen. Biochim. Biophys. Acta 1798, 2048-2057. doi: 10.1016/j.bbamem.2010.07.014
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2011). Förutspådd expansion av familjen claudin multigene. FEBS Lett. 585, 606–612. doi: 10.1016/j.febslet.2011.01.028
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Nenni, M. J., Fisher, M. E., James-Zorn, C., Pells, T. J., Ponferrada, V., Chu, S., et al. (2019). Xenbase: underlätta användningen av Xenopus för att modellera mänsklig sjukdom. Front. Physiol. 10:154. doi: 10.3389/fphys.2019.00154
PubMed Abstrakt / CrossRef fulltext / Google Scholar
det finns många olika sätt att hitta en produkt på. (2003). Storlek-selektiv lossning av blod-hjärnbarriären i claudin-5-bristfälliga möss. J. Cell Biol. 161, 653–660. doi: 10.1083 / jcb.200302070
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Ohtsuki, S., Sato, S., Yamaguchi, H., Kamoi, M., Asashima, T. och Terasaki, T. (2007). Exogent uttryck av claudin-5 inducerar barriäregenskaper i odlade råtta hjärnkapillära endotelceller. J. Cell Physiol. 210, 81–86. doi: 10.1002/jcp.20823
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Piontek, J., Fritzsche, S. Han är en av de mest kända i världen. (2011). Belysa principerna för den molekylära organisationen av heteropolymera snäva korsningssträngar. Cell Mol. Life Sci. 68, 3903–3918. doi: 10.1007 / s00018-011-0680-z
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Piontek, J., Winkler, L., Wolburg, H., Muller, S. L., Zuleger, N., Piehl, C., et al. (2008). Bildande av tät korsning: determinanter för homofil interaktion mellan klassiska claudiner. FASEB J. 22, 146-158. doi: 10.1096 / fj.07-8319com
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Radloff, J., Cornelius, V., Markov, A. G. och Amasheh, S. (2019). Caprate modulerar tarmbarriärfunktionen i porcine Peyers lapp follikelassocierat epitel. Int. J. Mol. Sci. 20:1418. doi: 10.3390 / Ijms20061418
PubMed Abstrakt | CrossRef fulltext | Google Scholar
Reinhold, A. K. och Rittner ,H. L. (2017). Barriärfunktion i perifera och centrala nervsystemet-en översyn. Pflug. Båge. EUR. J. Phys. 469, 123–134. doi: 10.1007/s00424-016-1920-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Resnick, M. B., Gavilanez, M., Newton, E., Konkin, T., Bhattacharya, B., Britt, D. E., et al. (2005). Claudin expression in gastric adenocarcinomas: a tissue microarray study with prognostic correlation. Hum. Pathol. 36, 886–892. doi: 10.1016/j.humpath.2005.05.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Rossa, J., Ploeger, C., Vorreiter, F., Saleh, T., Protze, J., Gunzel, D., et al. (2014). Claudin – 3 och Claudin-5-proteinvikning och montering i den täta korsningen styrs av icke-konserverade rester i transmembran 3 (TM3) och extracellulär slinga 2 (ECL2) segment. J. Biol. Chem. 289, 7641–7653. doi: 10.1074 / jbc.M113. 531012
PubMed Abstrakt / CrossRef fulltext / Google Scholar
det är en av de mest populära och mest populära spelen i världen. (2016). Genomutveckling i den allotetraploida grodan Xenopus laevis. Natur 538, 336-343.
Google Scholar
Soini, Y. (2011). Claudins i lungsjukdomar. Resp. Upp 12: 70.
Google Scholar
han är en av de mest kända och mest kända i världen. (2014). Kristallstruktur av en claudin ger inblick i arkitekturen av snäva korsningar. Vetenskap 344, 304-307. doi: 10.1126/vetenskap.1248571
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Tammaro, P., Shimomura, K. och Proks, P. (2009). ”Xenopus oocyter som ett heterologt uttryckssystem för att studera jonkanaler med patch-clamp-tekniken,” i kaliumkanaler: metoder och protokoll, ed. J. D. Lippiat (Totowa, NJ: Humana Press), 127-139.
Google Scholar
Tebbe, B., Mankertz, J., Schwarz, C., Amasheh, S., Fromm, M., Assaf, C., et al. (2002). Tight junction proteiner: en ny klass av integrerade membranproteiner-uttryck i human epidermis och i hacat keratinocyter. Arch Dermatol. Res 294, 14-18. doi: 10.1007 / s00403-001-0290-y
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Tomita, M., Hayashi, M. och Awazu, S. (1995). Absorptionsförbättrande mekanism för natriumkaprat och dekanoylkarnitin i Caco-2-celler. J. Pharmacol. Exp. Där. 272, 739–743.
Google Scholar
Turner, Jr (2000). ’Sätta pressningen’ på den snäva korsningen: förstå cytoskeletal reglering. Sperma. Cell Dev. Biol. 11, 301–308. doi: 10.1006/scdb.2000.0180
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Van Itallie, C. M. och Anderson, J. M. (2006). Claudiner och epitelial paracellulär transport. Annu. Rev. Physiol. 68, 403–429. doi: 10.1146/annurev.physiol.68.040104.131404
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2009). Kvantifierande krafter medierade av integrerade täta korsningsproteiner i cellcelladhesion. Exper. Mech. 49, 3–9. doi: 10.1007/s11340-007-9113-1
CrossRef Full Text | Google Scholar
Vitzthum, C., Stein, L., Brunner, N., Knittel, R., Fallier-Becker, P., and Amasheh, S. (2019). Xenopus oocytes as a heterologous expression system for analysis of tight junction proteins. FASEB J. 33, 5312–5319. doi: 10.1096/fj.201801451rr
PubMed Abstract | CrossRef Full Text | Google Scholar
Wilcox, E. R., Burton, Q. L., Naz, S., Riazuddin, S., Smith, T. N., Ploplis, B., et al. (2001). Mutationer i genen som kodar för tight junction claudin-14 orsakar autosomal recessiv dövhet DFNB29. Cell 104, 165-172. doi: 10.1016 / s0092-8674(01)00200-8
CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2003). Lokalisering av claudin – 3 i täta korsningar av blod-hjärnbarriären förloras selektivt under experimentell autoimmun encefalomyelit och humant glioblastom multiforme. Acta Neuropathol. 105, 586–592. doi: 10.1007 / s00401-003-0688-z
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Wylie, C. C., Brown, D., Godsave, S. F., Quarmby, J. och Heasman, J. (1985). Cytoskelettet av Xenopus oocyter och dess roll i utvecklingen. J. Embryol. Exp. Förvandla. 89, 1–15.
Google Scholar