annonser:
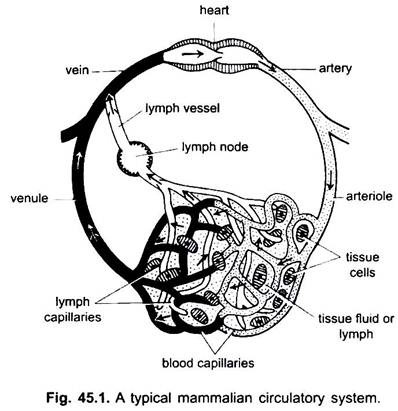
i cirkulationssystemet hos ryggradsdjur finns två system med detaljerade förgreningsrör, som ramify i hela kroppen och bär vätskor till vävnaderna. De är ett blodvaskulärt system och ett lymfsystem. När termen cirkulationssystem används, hänvisar det bara till blodvaskulärsystemet.
blodkärlsystemet är ett slutet system hos ryggradsdjur; det har ett kontraktilt hjärta och kontinuerliga rör som kallas kärl. Lymfsystemet är ett öppet system med lymfutrymmen. Blodkärlen som transporterar blod bort från hjärtat är artärer, som delar sig i tunnare arterioler, förgrenar sig till extremt tunna och små kapillärer.
en kapillärvägg är gjord av ett enda lager av tasselaterade endotelceller. Varje blodkärl, inklusive hjärtat, har en foder av endotelceller eller endotel. Blod kommer inte i kontakt direkt med vävnadsceller.
annonser:
ämnen passerar från och in i kapillärer genom vävnadsvätskan som finns i vävnadsutrymmen mellan celler. Utbyte av ämnen mellan kroppens blod och vävnadsceller sker genom kapillärväggarna. Detta orsakas av pinocytos i kapillärernas endotelceller.
under ett elektronmikroskop visar endotelcellerna många små vesiklar som är invaginationer av plasmamembranen. Dessa vesiklar rör sig från ena sidan av cellen och släpper ut innehållet på motsatt sida.
således ger blodet upp syre, näringsämne och hormoner till cellerna för metabolism, och cellerna ger ut vatten, koldioxid och kväveavfall i blodet för att bära dem till utsöndringsorganen för snabb eliminering.
kapillärerna bildar ett nätverk i alla kroppsvävnader utom brosk och epitel. Från kapillärerna passerar blodet in i tunna venuler som kombineras för att bilda vener som bär blod mot hjärtat. Men vissa vener (portalvener, njurvener och levervener) har kapillärer som är precis som artärer. Men allt blod passerar inte genom kapillärer i venuler.
annonser:
det finns också några genom kanaler mellan arterioler och venoler som ligger i vissa organ, såsom huden. Det finns också arteriovenösa anastomoser mellan arterioler och venoler som finns i siffrorna. Funktionen hos sådana anslutningar är inte klart känd, även om det hävdas att de reglerar blodtryck och cirkulation.
på vissa ställen sker utbyte av material mellan vävnader och blod genom tunnväggiga utrymmen eller sinusoider. I vissa organ kan ett blodkärl bilda ett lindat nätverk av små blodkärl som kallas rete mirabile (njurar, luftblåsa).
delar av cirkulationssystemet:
annonser:
i ackordater är cirkulationssystemet av sluten typ. Denna typ finns också i annelider (ryggradslösa djur). En annan typ är av öppen typ där kapillärer inte hittas. Det finns i blötdjur och leddjur. Här strömmar blodet genom artärer i olika organ, som passerar genom blodutrymmen eller bihålorna och sedan igen in i kärl (vener) till hjärtat.
cirkulationssystemet inkluderar hjärtat, artärerna, venerna, kapillärerna och blodet. Hjärta är ett modifierat blodkärl med muskelväggar, som periodiskt kontraherar för att pumpa blodet i olika delar av kroppen genom bestämda kärl.
artärer och deras grenar bildar ett arteriellt system som bär blodet från hjärtat. Vener och deras bifloder utgör ett venöst system som tar blodet från kapillärerna i artärer eller arterioler och bär det till hjärtat.
Portalsystem:
i portalsystemet returneras inte blodet direkt till hjärtat, men det finns ett interposerande organ (lever eller njure) under det återvändande blodet. Venen som bringar blod börjar i kapillärer och slutar i kapillärer, den berörda venen verkar både som afferent och efferent kärl, de afferenta kärlen slutar i kapillärer precis som artärer, då samlas blodet in i systemiska vener.
alla ryggradsdjur har ett leverportalsystem där blod passerar in i två uppsättningar kapillärer i levern. Lägre ryggradsdjur och embryon från högre ryggradsdjur har också ett njurportalsystem där blod passerar genom två uppsättningar kapillärer i njurarna innan de når hjärtat. Kapillärerna i hypofysen bildar ett hypofysportalsystem som är ett litet men ett viktigt system.
lymfsystemet:
det finns i ackordater utom cyklostomer och broskfiskar. Det inkluderar lymf-och lymfkärl. Lymf är en vävnadsvätska som finns bland kroppens celler. Det är blodplasma minus röda blodkroppar och vissa proteiner.
lymfkapillärer som bildar ett nätverk av tunna blindändande kärl som samlar lymf. Lymfkärl är tunnväggiga kärl som bildas av föreningen av blodkapillärer. Dessa tomma i ådror. Lymfkörtlar finns på lymfkärl hos däggdjur. Dessa bildar lymfocyter av blod som används för kroppsförsvar mot sjukdomar.
utveckling av hjärtat hos ryggradsdjur:
annonser:
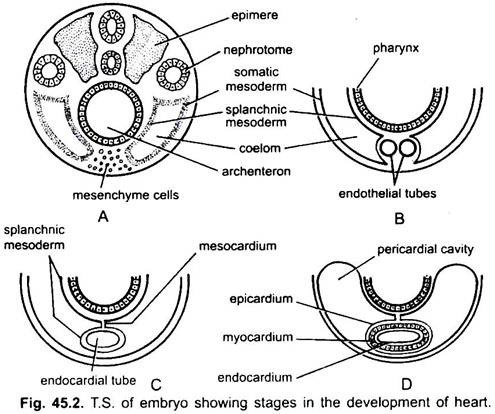
hjärtat är ett oparat organ men dess ursprung är bilateralt. I ett embryo bildar mesenkymet en grupp endokardiella celler under struphuvudet. Dessa celler blir ordnade för att bilda ett par tunna endotelrör. De två endotelrören smälter snart samman för att bilda ett enda endokardiellt rör som ligger i längdriktningen under svalget.
den splanchnic mesoderm som ligger under endodermen viks i längdriktningen runt endokardialröret. Detta tvåskiktsrör kommer att bilda hjärtat där splanchnic mesoderm tjocknar för att bilda en hjärt-eller muskelvägg i hjärtat och ett yttre tunt epikardium eller visceralt perikardium. Det endokardiella röret blir slemhinnan i hjärtat som kallas endokardium.
veck av splanchnic mesoderm möts ovan för att bilda ett dorsalt mesokardium som suspenderar hjärtat i coelom. Snart bildas en tvärgående septum bakom hjärtat som delar coelom i två kamrar, en främre perikardialhålighet som omsluter hjärtat och en bakre bukhålighet.
hjärtat är ett rakt rör men det ökar i längd och blir S-format eftersom dess ändar är fixerade. Utseende av ventiler, förträngning, skiljeväggar i hjärtat och differentiella förtjockningar av dess väggar bildar tre eller fyra kamrar i hjärtat.
1. Single-Chambered hjärta:
i Amphioxus (primitivt ackordat) finns inte ett sant hjärta. En del av ventral aorta under svalget är muskulös och kontraktil och fungerar som hjärta.
2. Två Kammare Hjärta:
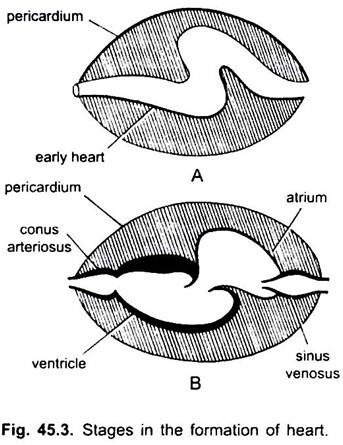
i cyklostomer finns fyra kamrar anordnade i linjär ordning-en tunnväggig sinus venosus, ett något muskulärt atrium (auricle), en muskulär ventrikel och en muskulär Conus arteriosus eller bulbus cordis. Det ligger i kroppshålan där andra viscerala organ också är närvarande.
av fyra kamrar motsvarar endast atrium och ventrikel de fyra kamrarna (Parade atria och parade ventriklar) hos de högre ryggradsdjuren. I hjärtats utveckling har många förändringar skett.
Elasmobranchs:
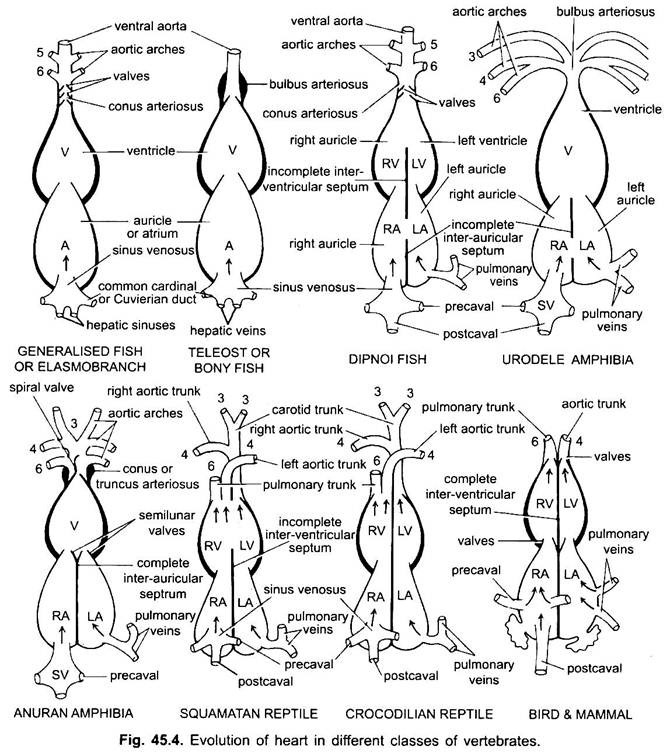
förutom Dipnoi, cirkulationssystemet i fiskar från cyklostomer till teleost, går endast ooxiderat blod till hjärtat, därifrån pumpas det till gälarna, luftas och distribueras sedan till kroppen. Hjärtat av broskig hundfisk är muskulärt och dorsoventralt böjt S-format rör med fyra fack i en linjär serie.
de är sinus venosus och atrium för att ta emot venöst blod och en ventrikel och conus arteriosus för att pumpa detta blod. Hjärtat är ett grenigt venöst hjärta. Sinus venosus och conus arteriosus är tillbehörskammare. Atrium och ventrikel är sanna kamrar, så det är ett 2-kammarhjärta.
sinus venosus öppnar främre in i atrium genom sinu-atriell öppning bevakad av ett par ventiler. Atrium ligger dorsalt till ventrikel och öppnar ventralt in i ventrikeln genom en atrio-ventrikulär öppning bevakad av ett par ventiler. Den tjockväggiga, muskulösa ventrikeln öppnar sig i en smal Conus arteriosus innehållande ventiler i två serier.
hjärtat är inneslutet i perikardialhålan separerad från kroppshålan med en tvärgående septum. Conus pierces perikardiet och blir kontinuerligt med ventral aorta. Perikardialhålan kommunicerar med kroppshålan genom två perforeringar i tvärgående septum.
Teleost:
deras hjärta liknar clasmobranchs. I teleosts reduceras konusen och har ett enda par ventiler. Den proximala delen av ventral aorta nära conus blir kraftigt förstorad och tjockväggig, kallad bulbus arteriosus. Det är elastiskt och dilaterar vid tidpunkten för ventrikulär sammandragning. Hjärtat är sålunda 2-kammare med en enda blodcirkulation.
3. Trekammat hjärta:
Dipnoi:
i difoer delar en septum atriumet i en höger och vänster kammare. Detta är korrelerat med användningen av simblåsan som ett andningsorgan och representerar det första steget mot utvecklingen av dubbeltypets cirkulationssystem där både syresatt och ooxygenerat blod kommer in i hjärtat och hålls åtskilda.
blod från lungfiskens högra aurikel passerar in i höger kammare och pumpas sedan in i den primitiva lungliknande gasblåsan av lungartärer som förgrenar sig från det sjätte paret av aortabågar. Det syresatta blodet återvänder till vänster atrium genom lungvener som amfibier.
Amfibia:
i amfibier skiftar dorsal atrium anterior till ventrikel. Sinus venosus öppnar sig i höger atrium dorsalt och inte bakom. Atriumet är helt uppdelat i höger och vänster kammare och har ingen foramen ovale i det inter-aurikulära septumet, som förblir öppet i dipnoans.
djupa fickor utvecklas i ventrikulärhålan. Conus arteriosus delar sig i systemiska och lungkärl med en spiralventil. I lunga mindre salamandrar är det inter-atriella septumet ofullständigt och lungorna är frånvarande.
Reptilia:
i reptiler är hjärtat vidare avancerat. Atriumet är alltid helt separerat i en höger och vänster kammare, och i många former är sinus venosus införlivad i väggen i det högra atriumet. Ventrikeln är också delvis uppdelad av en septum i de flesta reptiler, och i alligatorerna och krokodilerna är helt tvåkammare.
detta innebär att syresatt blod som kommer från lungorna till vänster sida av hjärtat i huvudsak separeras från det icke-syresatt blod från kroppen till höger sida. Således är de två blodtyperna i krokodilier helt separerade och nästan fullständiga i andra reptiler, men viss blandning förekommer i andra delar av cirkulationssystemet.
den embryonala Conus arteriosus delas upp i tre istället för två kärl:
(i) Lungbåge som bär blod till lungorna från höger sida av ventrikeln.
(ii) höger systemisk aorta som transporterar blod från vänster sida av ventrikeln till kroppen genom höger fjärde aortabåge.
(iii) vänster systemisk kommer från höger kammare till vänster fjärde aortabågen.
vid kontaktpunkten med den systemiska aortan från vänster ventrikel, även hos krokodiler, finns en öppning mellan de två, kallad foramen av Panizzae där det kan finnas viss blandning av de två blodtyperna. Således representerar reptilianhjärta övergångshjärtat mot amfibiehjärta-2 kompletta auriklar och 2 ofullständiga ventriklar med lite blandning av blod i höger och vänster systemisk.
4. Fyra kammare hjärta:
Aves och Mammalia:
hos fåglar är ventrikeln helt uppdelad i två, så att hjärtat är fyra kammare (2 auriklar och 2 ventriklar). Det finns fullständig separation av venöst och arteriellt blod. Den systemiska aortan lämnar vänster ventrikel och bär blod till huvudet och kroppen. Medan lungartären lämnar höger kammare och bär blod till lungorna för syresättning.
således finns det Dubbel cirkulation där det inte finns någon blandning av blod på någon plats. Sinus venosus är helt införlivad i höger auricle, som tar emot två precavals och en postcaval. Den vänstra aurikeln tar emot syresatt blod genom lungorna, conus arteriosus är frånvarande, lungortan uppstår från höger kammare och enstaka systemisk aorta uppstår från vänster ventrikel, och båda har ventiler vid sina baser.
modifieringar av Aortabågar hos ryggradsdjur:
embryonala artärer:
när hjärtat bildas i ett ryggradsembryo uppträder ett blodkärl som kallas ventral aorta mitt i ventralt under svalget, vilket snart blir anslutet till conus arteriosus. Den ventrala aortan som härrör från hjärtat löper framåt under struphuvudet och delar sig framåt i ett par yttre halspulsåder i huvudet.
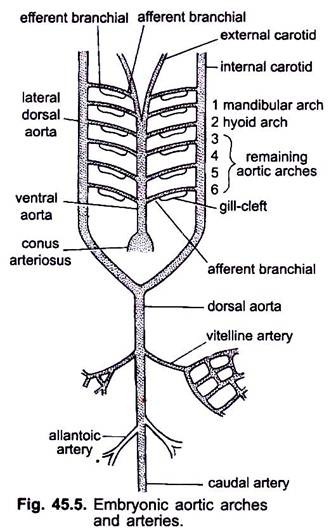
den ventrala aortan avger 6 par laterala aortabågar på lika avstånd som löper genom de viscerala bågarna. Varje aortabåge har en ventral afferent grenartär som bär venöst blod till gälen och en dorsal efferent grenartär som tar syresatt blod från gälen. De efferenta grenartärerna på vardera sidan förenas dorsalt med lateral dorsal aorta eller radix aorta som kommer in i huvudet som inre halspulsåder.
den första aortabågen är en mandibulär aortabåge, den andra är en hyoid aortabåge, de återstående kallas tredje, fjärde, femte och sjätte aortabågen. Den laterala dossala aortan smälter bakom svalget för att bilda en dorsal aorta som fortsätter mitt i dorsalt i svansen som en kaudal artär.
från dorsal aorta uppstår Parade och oparade artärer som levererar olika organ i kroppen. I ett embryo med en äggula säck uppstår ett par vitellinartärer från dorsal aorta och levererar äggula. I embryon av amnioter uppstår ett par navel-eller allantoiska artärer från dorsal aorta som levererar blod till allantois.
hos en vuxen smälter vitellinartärerna för att bilda den huvudsakliga mesenteriska artären, huvuddelen av de allantoiska artärerna förloras, men deras rester bildar hypogastriska eller inre iliacartärer.
Aortabågar hos ryggradsdjur:
i olika vuxna ryggradsdjur verkar artärsystemet vara annorlunda, men de är byggda på samma grundläggande grundläggande plan. Skillnaden beror på ökad komplexitet i hjärtat på grund av en förändring i andning från gälar till lungor. Det finns en progressiv minskning av antalet aortabågar i ryggradsserien.
Cyclostomata:
i Petromyzon finns det 7 par aortabågar. I andra cyklostomer varierar dessa från 6 par i Myxin och 15 Par i Eptatretus.
Fiskarna:
även om sex anses ha varit det grundläggande antalet aortabågar för fiskar. Detta antal reduceras till fem även i hajar och strålar med förlusten av det första paret, den mandibulära aortabågen eller den representeras av en efferent pseudobranchial artär. I de flesta Benfiskar försvinner både mandibulära (i) och hyoid (ii) aortabågar eller reduceras mycket.
i Polypterus och Dipnoi (lungfiskar) är gälarna inte väl utvecklade. Så lungartären uppstår från den efferenta delen av 6: e bågen på vardera sidan som levererar blod till luftblåsan eller lungan. I elasmobranchs och Dipnoi har varje båge en afferent och två efferenta grenartärer (bildade genom splittring) i varje gill. I Benfiskar har varje gill en afferent och en efferent artär.
i tetrapoda bryts inte aortabågar upp i afferenta och efferenta delar eftersom sanna inre gälar saknas. I alla tetrapoda försvinner de första och andra bågarna.
Amfibia:
här visar aortabågarna modifiering på grund av förlust av gälar och utseende av lungorna. I urodeler finns det yttre gälar som andningsorgan förutom lungor. III, IV, V och VI aortabågar är närvarande, även om det femte paret är mycket reducerat I Siren, Amphiuma och Necturus. Aortabågarna bryts inte i de yttre gälarna i afferenta och efferenta delar, eftersom grenar som härrör från IV -, V-och VI-aortabågar bildar kapillärer i de yttre gälarna.
den laterala dorsala aortaen mellan III-och IV-aortabågarna kvarstår som en vaskulär anslutning, ductus caroticus. VI aortabågen bildar den pulmo-kutana bågen eller artären på vardera sidan och tar blod till lungan och huden. Det behåller också en förbindelse med den laterala dorsala aortan som kallas en ductus arteriosus (kanal av Botalli).
i larven av anuran (groda tadpole) är Arrangemang av aortabågar som en vuxen urodele på grund av närvaron av gälar. Vid metamorfos, med förlust av gälar, försvinner i, II och V aortabågar helt, endast IIIrd, IVth och Vlth aortabågar är närvarande. Den laterala dorsala aortan mellan den tredje och fjärde aortabågen (ductus caroticus) försvinner också. Således blir den tredje aortabågen tillsammans med en del av den ventrala aortan carotidbågen som bär syresatt blod till huvudregionen.
den fjärde aortabågen längs sin laterala dorsala aorta bildar den systemiska bågen. Den sjätte aortabågen blir den pulmokutana bågen som levererar venöst blod till lungor och hud. Ductus arteriosus försvinner under metamorfos. Således har vuxna anuraner endast III, IV och VI aortabågar. Dessa behålls också av amnioter.
Reptilia:
i reptiler ersätts gälarna helt av lungor. Endast III, IV och VI aortabågar är närvarande. Med partiell separation av ventrikeln i två delar delas den distala delen av conus arteriosus och hela ventral aorta i tre kärl, dvs två aorta eller systemiska och en lung. Höger systemisk båge (IV) uppstår från vänster ventrikel som transporterar syresatt blod till karotidbågen (III). Den vänstra systemiska (IV) och lungortae (VI) har sitt ursprung från höger kammare. Den vänstra systemiska bär deoxygenerat eller blandat blod till kroppen genom dorsal aorta.
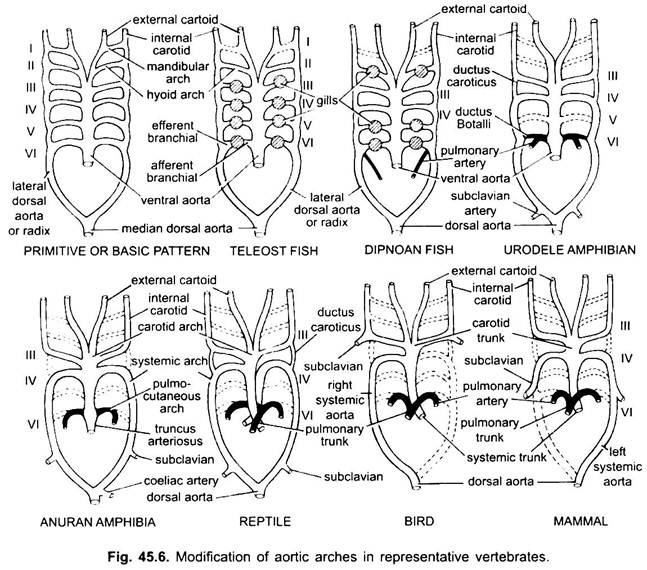
medan lungartären tar deoxygenerat blod till lungorna. Ductus caroticus försvinner, men den kvarstår i ormar och några ödlor (Uromastix). Ductus arteriosus försvinner i de flesta reptiler även om det kvarstår i en reducerad form i Sphenodon och vissa sköldpaddor. På grund av blandning av blod är reptiler kallblodiga djur som fiskar och amfibier.
Aves:
hos fåglar är III, IV och VI aortabågar närvarande. De följer det allmänna mönstret av reptiler med vissa skillnader. Med den fullständiga uppdelningen av ventrikeln i två delar har conus arteriosus och ventral aorta delats för att bilda två kärl, systemisk aorta som härrör från vänster ventrikel och en pulmonell aorta från höger ventrikel.
tredje aortabågen med rester av laterala och ventrala aorta bildar karotiderna som uppstår från systemisk aorta. Fjärde aortabågen bildar den systemiska aortan endast på höger sida. Den förenar sig med den laterala aortan på sin egen sida och bildar dorsal aorta. En del av den fjärde aortabågen på vänster sida bildar den vänstra subklaviska artären, resten tillsammans med dess laterala dorsala aorta försvinner. Den sjätte aortabågen bildar lungorta. Ductus caroticus och ductus arteriosus försvinner.
Mammalia:
hos däggdjur kvarstår även III, IV och vi aoriska bågar. Ventrikeln är helt uppdelad i två delar.
conus arteriosus och ventral aorta delas för att bilda två kärl:
(i) en systemisk aorta som härrör från vänster ventrikel och
(ii) en pulmonell aorta från höger ventrikel. Tredje aortabågen med rester av laterala och ventrala aorta bildar karotidbågen.
fjärde aortabågen bildar den systemiska aortan endast på vänster sida, medan på höger sida bildar dess proximala del en innominat och höger subklavisk artär, försvinner resten tillsammans med dess laterala dorsala aorta. Sjätte aortabågen bildar lungorta. Ductus arteriosus degenererar men det kvarstår i vissa tills kläckning eller födelse i reducerad form på vänster sida som en tunn ligamentum arteriosum.
Venöst System:
Embryonala Vener:
i alla ryggradsembryon är vensystemet enkelt och liknande. Venerna är mestadels Parade och symmetriskt anordnade. I embryon utan äggula säck bildas en sub – tarmven i splanchnic mesoderm under tarmen.
det slingrar runt anus och fortsätter bakåt som:
(i) en kaudal ven i svansen. I alla embryon som har en äggula (oavsett om de innehåller äggula eller inte) uppstår ett par vitellinvener från äggula och förenar den bakre delen av det utvecklande hjärtat som blir sinus venosus. I själva verket är fusionen av dessa vitellina vener ansvarig för hjärtbildning i Benfiskar, reptiler och fåglar. Varje vitellinven i sin bakre ände förenar den subintestinala venen som bildas på samma sätt som i embryona som inte har någon äggula.
(ii) ett par subkardinala vener uppstår mellan njurarna och förenar den kaudala venen.
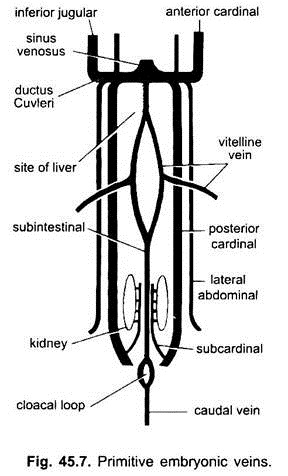
(iii) Parade främre och bakre kardinalvener bildas, vilka bär blodet från respektive huvud och bakre delar av kroppen. De främre och bakre kardinalvenerna på varje sida förenas för att bilda en ductus Cuvieri eller vanlig kardinalven som passerar inåt genom det tvärgående septumet för att komma in i sinus venosus.
hos fiskar och salamandrar (urodeles) kommer en underlägsen halsven från den ventrala sidan av huvudet för att gå med i den gemensamma kardinalvenen. Den har ingen homolog hos andra ryggradsdjur. I en amnioter kommer ett par laterala eller ventrala bukvener från kroppsväggen för att komma in i de gemensamma kardinalvenerna.
Fiskarna:
vanlig kardinalven (kanal av Cuvier) kommer in i sinus venosus från varje sida och bildas genom fusion av främre och bakre kardinaler. Blod från huvudet samlas in av de främre kardinalerna, och blod från njurarna och gonaderna samlas in av de bakre kardinalerna.
även in i kanalerna i Cuvier är de parade laterala bukvenerna som tar emot blod från kroppsväggen och parade bilagor. Njurportalsystemet består av den kaudala venen och två njurportalvener belägna i sidled till njurarna som kapillariserades i njurarna.
Leverportalsystemet bär blod från magen och tarmen och returnerar det till levern, där det, efter att ha passerat genom serier av sinusoider, kommer in i sinus venosus genom Parade hapatiska vener.
i teleosts saknas de laterala bukvenerna så att blod från subklaverna, som dränerar pectoral bihang, kommer in i sinus venosus direkt, och blodet från iliac vener, som dränerar bäckenbilagorna, passerar in i postkardinalerna.
i dipnoans är en enda ventral bukven närvarande, förmodligen härledd från en fusion av laterala buken. Denna ven tar emot blod från ILAC: erna genom Parade bäckenvener och går in i Cuviers högra kanal. Vidare, från det högra postkardinala systemet, en ny ven, gör postclaval sitt utseende som är av stor betydelse hos högre ryggradsdjur. Den är ansluten till den kaudala venen och passerar framåt genom levern till sinus.
i embryon av amnioter är de laterala bukvenerna kända som navel-eller allantoiska vener eftersom de dränerar allantois. Vitellinen och navelsträngarna förloras vid födseln, bara deras rester kvarstår.
ändringar av vener:
som i fallet med artärer är venerna hos olika ryggradsdjur ordnade på samma grundläggande plan. Eventuella variationer som visas av dem följer en sekvens i vertebratserien. I sin utveckling passerar det venösa systemet av högre ryggradsdjur genom de steg som ses i embryon av lägre former.
1. När levern bildas den proximala delen av vitellinvenerna eller subintestinala venerna bildar levervenerna mellan levern och hjärtat, den distala delen av den vänstra vitellinvenen eller subintestinala venen blir leverportalvenen som bildar sinusoider i levern för att ge upphov till ett leverportalsystem som finns i alla.
2. Främre kardinalvener kvarstår som inre halsvener.
3. Förutom i fiskar blir de vanliga kardinalvenerna de precavala venerna som kommer in i sinus venosus (amfibier, reptiler) eller höger auricle (fåglar, däggdjur). En subklavisk ven bildas i varje framben som förenar den prekavala venen.
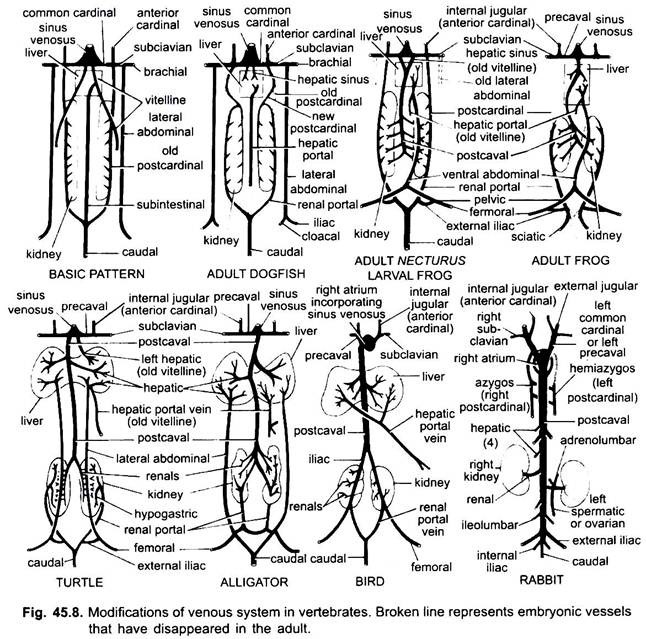
4. Den kaudala venen förlorar sina förbindelser med subintestinala och subkardinala vener, dess främre del delas upp i två grenar som förenar de bakre kardinalvenerna i alla utom däggdjur.
5. Posterior kardinal vener kvarstår som sådan i fiskar, men i andra varje bryts upp i två delar, en främre del som försvinner i amfibier, reptiler, och fåglar, men hos däggdjur den högra främre delen bildar en azygos ven, den vänstra kan bilda en hemiazygos ven som förlorar sin förbindelse med precaval och förenar azygos ven av en tvärgående anastomos; de bakre delarna av de bakre kardinal vener som har anslutit sig till stjärtfenan ven, blir njurportal vener.
njurportalvenen på varje sida bildar kapillärer i njurarna för att bilda ett njurportalsystem som är komplett i fiskar, amfibier och reptiler, men blir mycket reducerad hos fåglar och saknas hos däggdjur eftersom de bakre delarna av bakre kardinalvener försvinner hos däggdjur.
6. Fiskens njurportalår samlar bara blod från svansen, men i amfibier, reptiler och fåglar ökar de också venerna från benen.
7. I vissa fiskar och alla tetrapoda går vitellinvenerna i subkardinalvenen för att bilda en postkavalven. I amfibier och reptiler sträcker sig postkavalvenen till njurarnas bakre ändar, hos fåglar förenar den njurportalvenerna, vilket minskar njurportalsystemet, hos däggdjur förenar postkavalvenen venerna från benen och svansen, så att njurportalsystemet elimineras helt.
8. De två laterala bukvenerna kvarstår som sådana hos fiskar, men från Dipnoi uppåt smälter de samman för att bilda en främre bukven som förenar leverportalvenen nära levern, den främre bukvenen förbinder således njurportalen och leverportalsystemen. En iliac ven bildas i varje bakben som förenar den främre bukvenen. Den främre bukvenen börjar förlora sin betydelse i reptiler, hos fåglar modifieras den för att bilda de epigastriska och coccygeo-mesenteriska venerna, hos däggdjur försvinner den, utom i Tachyglossus.
9. I luftandningsformer växer lungvenerna ut från vänster öron och går in i lungorna.
10. Embryonets navel (allantoiska) vener försvinner när lungorna blir funktionella som andningsorgan.
Tetrapoda:
amfibiernas venösa system liknar mycket lungfiskernas, förutom att bukvenen kommer in i leverportalsystemet snarare än sinus venosus. Främre kardinaler kvarstår som inre halsvener hos alla vuxna tetrapoder. Inferior jugular vener är frånvarande. Vanliga kardinaler blir den främre vena cavae eller precavals, som går med sinus venosus i amfibier och reptiler.
hos fåglar och däggdjur kommer precavals direkt in i hjärtans högra aurikel, eftersom sinus venosus saknas. Hos människa, katt, etc., vänster precaval förloras så blodet på vänster sida kommer in i höger precaval genom en brachio-cephalisk gren. I lunglösa salamandrar är lungvenerna frånvarande eftersom inter-atrial septum är ofullständig.
post-kardinalvenerna förenar kaudalvenen bakåt och precavals framåt för att bilda de vanliga kardinalerna i larvgroda och Necturus.
i reptiler finns det större utveckling av lungvener och postkavalven och minskning av betydelsen av njurportalsystem som leder blod till njurarna från den bakre delen av kroppen.
hos fåglar finns det två funktionella prekavala vener som bildas av föreningen av jugular och subclavian på varje sida och en komplett postkaval. Det tar emot blod från lemmarna genom njurportalerna, som passerar genom njurarna, men bryter inte upp i kapillärer och är inte jämförbara med njurportalerna hos nedre ryggradsdjur.
hos däggdjur kan det finnas en eller två prekavala vener. Det finns en enda postcaval ven, vars embryologiska utveckling är mycket komplicerad. Alla kavalvenerna går direkt in i höger öron, eftersom sinus venosus absorberas i väggen i denna kammare i hjärtat i embryonalt liv. Det finns inget njurportalsystem även om leverportalsystemet mycket liknar det hos andra ryggradsdjur.