- Introducere
- rezultate
- șoferul DBH-Cre marchează și este limitat la regiunile noradrenergice care exprimă TH în trunchiul cerebral care sunt inhibate de administrarea CNO
- perturbarea mediată de CNO-hM4D a neuronilor noradrenergici la șoarecii adulți
- perturbarea CNO-hM4D a neuronilor DBH-Cre are ca rezultat un Deficit Hipercapnic
- controale obișnuite ale fraților injectați cu 10 mg / kg CNO au arătat un deficit Ventilatoric Hipercapnic
- animalele de tip sălbatic obișnuite și Neabituate injectate cu soluție salină nu au prezentat modificări Presalină și Postsalină
- raporturile concentrației clozapinei la CNO sunt mai mari în creier decât în ser
- discuție
- materiale și metode
- aprobare etică
- reproducerea, fondul Genetic și întreținerea șoarecilor
- Histologie
- electrofiziologie
- pletismografie
- cromatografie lichidă-spectrometrie de masă
- colectarea și analiza datelor
- pletismografie
- statistici
- pletismografie
- cromatografie lichidă-spectrometrie de masă
- declarație de etică
- contribuțiile autorului
- finanțare
- Declarație privind conflictul de interese
- mulțumiri
Introducere
rețelele neuronale care stau la baza reflexului chemosenzorial respirator sunt o țintă primară pentru înțelegerea etiologiei mai multor tulburări comportamentale și fiziologice. Reflexele chemosenzoriale perturbate se presupune că joacă un rol atât în tulburările congenitale, cât și în cele adulte, inclusiv sindromul morții subite a sugarului (SIDS), sindromul Central Congenital de hipoventilație (CCHS), apneea centrală a somnului, respirația anormală în sindromul Rett și sindromul de hipoventilație a obezității (Obonai și colab., 1998; Ozawa și colab., 2003; Amiel și colab., 2009; Ramanantsoa și colab., 2011; Lavezzi și colab., 2013). Sub ipoteza alarmei false de sufocare, se consideră că activarea chemosenzorială inadecvată sau hipersensibilitatea joacă un rol în subseturile de pacienți care suferă de tulburare de panică (Klein, 1993). Disfuncția chemosenzorială poate juca, de asemenea, un rol în bolile neurodegenerative prin respirația dezordonată a somnului, care este asociată cu progresia accelerată (Hakim și colab., 2012; Verbraecken și McNicholas, 2013; Bahia și Pereira, 2015; Snyder și colab., 2017). Astfel, o mai bună înțelegere a rețelelor chemosenzoriale ale trunchiului cerebral va oferi indicii importante pentru o serie de patologii comportamentale și fiziologice.
receptori de designer Farmacogenetic activi exclusiv de medicamente de designer (DREADDs) (Armbruster și colab., 2007) au fost angajați într-o serie de studii pentru a cartografia populațiile neuronale din chemoreflex respirator (răspunsul respirator la nivelurile crescute de CO2 din sânge). Tehnologia DREADD în combinație cu implementarea genetică intersecțională a fost utilizată pentru a reduce la tăcere populațiile neuronale foarte vizate, observând în același timp funcția respiratorie la șoarecii conștienți și neîngrădiți de laboratorul nostru și de alții, evitând multe confuzii din abordările anterioare de cartografiere a circuitelor (Ray și colab., 2011, 2013; Brust și colab., 2014; Hennessy și colab., 2017; soare și raze, 2017; soare și colab., 2017). Acestea și majoritatea celorlalte studii au inclus aproape întotdeauna CNO numai controale care nu exprimă DREADD care nu au prezentat efecte chemosenzoriale sau alte efecte respiratorii, argumentând că CNO nu a avut efecte în afara țintei asupra respirației la șoareci conștienți și neîngrădiți, în ciuda dozei mari utilizate, 10 mg/kg. Cu toate acestea, s-a constatat în alte câteva studii că CNO și produsele sale de metabolizare a spatelui ar putea avea efecte în afara țintei asupra comportamentului și locomoției într-o varietate de teste, dar ieșirea respiratorie nu a fost abordată (Guettier și colab., 2009; Joober, 2010; MacLaren și colab., 2016; Gomez și colab., 2017; Ilg și colab., 2018; Mahler și Aston-Jones, 2018; Manvich și colab., 2018; Padovan-Hernandez și Knackstedt, 2018). De asemenea, sa demonstrat că CNO și metaboliții săi nu sunt distribuiți echivalent între sistemul circulator și creier (Gomez și colab., 2017).
o a doua preocupare neadresată apare din stresul indus de paradigma experimentală, inclusiv deținerea animalelor într-o cameră de pletismografie barometrică a întregului corp, manipularea, injecția intra-peritoneală și măsurătorile temperaturii rectale. În studiile respiratorii anterioare DREADD, șoarecii na au fost introduși într-o cameră mică (140-400 mls) și li s-a permis să se aclimatizeze cu 20-40 de minute înainte de achiziția de date (Ray și colab., 2011, 2013; Brust și colab., 2014; Hennessy și colab., 2017; soare și raze, 2017; soare și colab., 2017). Cu toate acestea, nu este clar dacă această perioadă de timp este suficientă pentru a minimiza modificările respiratorii induse de stres care pot acționa ca un factor de interacțiune cu efectele clozapinei.
în studiile noastre pentru a examina rolul sistemului noradrenergic (NA) în controlul respirator, am căutat să folosim RC bine stabilit::Alela DREADD P_HM4D (Ray și colab., 2011) pentru a testa rolul neuronilor NA în respirația inițială și hipercapnică la animalele adulte nerestricționate folosind pletismografia întregului corp. Deoarece neuronii noradrenalinei și NA sunt, de asemenea, cunoscuți că joacă un rol central în răspunsurile la stres (Valentino și Van Bockstaele, 2008; Chen și colab., 2018), am efectuat o serie de studii pentru a compara obișnuința extinsă (expuneri multiple la paradigma experimentală prin care animalul învață că experiența nu amenință sau supraviețuiește) și ridicată vs. doze CNO scăzute (1 mg/kg) cu protocoale DREADD publicate anterior (Ray și colab., 2011, 2013; Brust și colab., 2014; Hennessy și colab., 2017; soare și raze, 2017; soare și colab., 2017). Aici, arătăm pentru prima dată că dozele sistemice mari de CNO sunt capabile să provoace efecte în afara țintei asupra unei funcții respiratorii autonome la șoarecii conștienți. De asemenea, Dezvăluim că efectul off-țintă al CNO asupra debitului respirator chemosenzorial este demascat efectiv de o obișnuință extinsă și, prin urmare, nu ar fi evident în studiile anterioare de control CNO care nu au obișnuit animalele înainte de măsurarea respiratorie, utilizând doar o perioadă scurtă de aclimatizare înainte de colectarea datelor. Împreună, aceste rezultate sugerează că populațiile neuronale cartografiate anterior pot afecta indirect controlul respirator prin roluri potențiale în reglarea răspunsurilor la stres. În special, aceste date se aliniază cu rapoartele recente care sugerează că CNO nu este inert biologic la doze mari prin conversia metabolică la clozapină (MacLaren și colab., 2016; Gomez și colab., 2017) și că efectele comportamentale în afara țintei se pot manifesta nu numai din perturbarea circuitelor comportamentale, ci și din perturbări ale circuitelor autonome subiacente și ale homeostaziei.
rezultate
șoferul DBH-Cre marchează și este limitat la regiunile noradrenergice care exprimă TH în trunchiul cerebral care sunt inhibate de administrarea CNO
pentru a examina expresia și specificitatea liniei DBH-Cre, am folosit o singură schemă de reproducere a recombinazei (figura 1A) în care am trecut șoferul la linia Ai9 (Madisen și colab., 2010), care exprimă un tdtomato floxat. Colorarea cu un anticorp tirozin hidroxilază (TH) a arătat că expresia tdTomato în trunchiul cerebral a fost limitată la regiunile care exprimă TH, inclusiv nucleele locus coeruleus, A5, a1, A2, A7, SubCV și SubCD așa cum era de așteptat (figura 1b). Pentru a confirma că neuronii NA care exprimă receptorul hM4D au fost receptivi la CNO, am efectuat înregistrări pe neuronii locus coeruleus (LC), unde am observat o inhibare a arderii neuronilor la aplicarea băii CNO (N = 3, figura 1C).
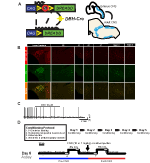
Figura 1. Protocoale schematice și respiratorii de reproducere. RC::Alela P_hM4D este combinată cu o linie de dopamină-beta-hidroxilază (DBH)-Cre pentru a obține expresia specifică celulară a receptorului hM4D în neuronii noradrenergici (NA) (a). Expresia fluorescentă a tdTomato în DBH-Cre; Ai9 secțiunile coronale ale creierului coexprimă cu tirozin-hidroxilază (TH) în nucleele na ale trunchiului cerebral (B). În înregistrarea slice a na locus coeruleus înainte și după aplicarea CNO, demonstrând funcționalitatea receptorului DREADD în neuronii vizați (C). Protocolul respirator. Șoarecii obișnuiți sunt supuși unui protocol obișnuit de 5 zile constând în manipulare, măsurarea temperaturii sondei rectale, injecție salină și expunerea la camera de pletismografie. În a șasea zi, atât șoarecii obișnuiți, cât și cei nehabituați, sub un test hipercapnic, în care animalul este plasat în camera respiratorie și lăsat să se aclimatizeze în condițiile de bază ale aerului din cameră. Animalul este apoi expus la 20 min de 5% CO2 urmat de 20 min de aer din cameră. Animalul este apoi injectat intraperitoneal cu CNO sau soluție salină, urmat de încă 20 de minute de aer post-injecție, 20 de minute de 5% CO2 și 20 de minute de aer din cameră. Casetele deschise delimitează timpii de colectare a datelor (D).
perturbarea mediată de CNO-hM4D a neuronilor noradrenergici la șoarecii adulți
pentru a examina rolul neuronilor NA sub respirația inițială și hipercapnică, am folosit sistemul DREADD inhibitor RC:P_hM4D încrucișat cu driverul DBH-Cre. Folosind pletismografia întregului corp (Ray și colab., 2011), am măsurat răspunsurile ventilatorii ale animalelor adulte nerestricționate în condiții de aer din cameră (21% O2/79% N2) și hipercapnic (5% CO2/21% O2/74% N2) înainte și după administrarea CNO (figura 1D). Pentru a aborda dozarea CNO și stresul potențial indus de proiectul nostru experimental, animalele au fost supuse uneia dintre cele patru condiții: (1) nehabituate și injectate cu 10 mg/kg CNO; (2) obișnuite și injectate cu 10 mg/kg CNO; (3) nehabituate și injectate cu 1 mg/kg CNO; sau (4) obișnuite și injectate cu 1 mg/kg CNO. Obișnuința a constat într-un proces de 5 zile care implică manipularea, sonda de temperatură rectală, injecția salină și expunerea la camera de pletismografie în fiecare zi timp de 30 de minute, în timp ce animalele na au fost expuse doar la o perioadă de aclimatizare a camerei de 20-45 de minute imediat înainte de colectarea datelor, așa cum s-a făcut în studiile anterioare (Ray și colab., 2011, 2013; Brust și colab., 2014; Hennessy și colab., 2017; soare și raze, 2017; soare și colab., 2017). Parametrii respiratori măsurați au inclus rata respiratorie RR, volumul mareelor (VT), ventilația minusculă (VE), consumul de oxigen (VO2), ventilația minusculă normalizată la consumul de oxigen (VE/VO2), frecvența apneei, frecvența suspinului și coeficienții de variație pentru intervalul și amplitudinea interbreath (instabilitate periodică și de volum). Ca un control suplimentar, am comparat, de asemenea, animalele de tip sălbatic obișnuite și neabituate injectate cu soluție salină.
perturbarea CNO-hM4D a neuronilor DBH-Cre are ca rezultat un Deficit Hipercapnic
în trei dintre condiții, DBH-Cre; RC::Animalele P_hM4D au prezentat o reducere a VE și VE/VO2 după administrarea CNO, în timp ce controalele frate nu au arătat diferențe în valorile pre – și post-CNO. Animalele nehabituate injectate cu 10 mg/kg CNO au prezentat o reducere semnificativă a RR (-12,17%, p = 0,034) și VT (-30,87%, p = 0,0016), rezultând o reducere a VE (-38,64%, p = 0,0031) și o ușoară reducere a VO2 (-14,25%, p = 0,042) (Figura 2). Reducerea VE a fost mai mare decât scăderea VO2, rezultând o reducere globală a VE/VO2 (-26,89%, p = 0,0095). Animalele nehabituate injectate cu 1 mg/kg CNO au prezentat o tendință spre reducerea RR (-12,88%, p = 0,066) și reducerea semnificativă a VT (-16,52%, p = 0,00085) și VE (-28,08%, p = 0,0070), ducând la o reducere globală a VE/VO2 (-22,23%, p = 0,016) (Figura 3). În cele din urmă, animalele obișnuite injectate cu 1 mg/kg de CNO au prezentat o RR redusă (-10,77%, p = 0,074), tendință spre VE redusă (-25,07%, p = 0,074) și ve/VO2 global redus (-23,70%, p = 0,024) (Figura 4). Nu s-au observat modificări semnificative ale frecvenței apneei, frecvenței suspinului, instabilității periodice sau de volum sau temperaturii în nicio cohortă.
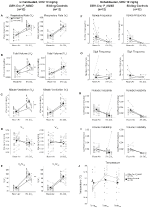
Figura 2. Unhabituated DBH-Cre; RC::p_hm4d animalele administrate 10 mg/kg de CNO prezintă un deficit hipercapnic, în timp ce controalele fraților nu prezintă nicio modificare. După administrarea CNO, DBH-Cre; RC::Animalele P_hM4D nu prezintă nicio modificare a ventilației aerului din încăpere și reduceri ale RR (A), VT (B), VE (C), VO2 (D) și VE/VO2 (e) în condiții hipercapnice, fără modificări ale frecvenței apneei (F), frecvenței suspinului (G), instabilității periodice (H) sau a volumului (I) și temperaturii (J). Controalele fraților nu au arătat nicio diferență în toate valorile pre și post – CNO.
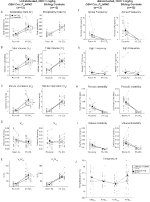
Figura 3. Animalele nehabituate DBH-Cre; RC::p_hm4d administrate 1 mg/kg de CNO prezintă un deficit hipercapnic, în timp ce controalele fraților nu prezintă nicio modificare. După administrarea CNO, DBH-Cre; RC::Animalele P_hM4D nu prezintă modificări semnificative ale ventilației aerului din cameră sau RR (A) și au prezentat reduceri ale VT (B), VE (C) și ve/VO2 (e) în condiții hipercapnice, fără modificări ale VO2 (D), frecvenței apneei (F), frecvenței suspinului (G), instabilității periodice (H) sau a volumului (I) și temperaturii (J). Controalele fraților nu au arătat nicio diferență în toate valorile pre și post – CNO.
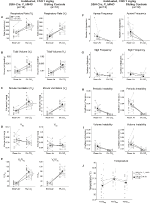
Figura 4. DBH-Cre obișnuit; RC:: p_hm4d animalele administrate 1 mg/kg de CNO prezintă un deficit hipercapnic, în timp ce controalele fraților nu prezintă nicio modificare. După administrarea CNO, DBH-Cre; RC:: animalele P_hM4D nu prezintă nicio modificare a ventilației aerului din încăpere și o reducere a RR (A) și VE/VO2 (e) în condiții hipercapnice,fără modificări semnificative ale VT (b),VE (C), VO2 (D), frecvența apneei (F), frecvența suspinului (G), instabilitatea periodică (H) sau a volumului (I) și temperatura (J). Controalele fraților nu au arătat nicio diferență în toate valorile pre și post – CNO.
controale obișnuite ale fraților injectați cu 10 mg / kg CNO au arătat un deficit Ventilatoric Hipercapnic
în ambele DBH-Cre; RC::P_hM4D și animalele de control habituate frate injectate cu 10 mg/kg de CNO, am observat o reducere semnificativă a VE / VO2 (p = 0,0235) mediată de scăderi ale RR (p = 0,00036) și VE (p = 0,037) (Figura 5). Cu toate acestea, spre deosebire de celelalte cohorte, nu a existat nicio diferență între DBH-Cre; RC::P_hM4D și animalele de control frate în acești parametri: RR (-12,72 vs.-10,22%, p = 0,6268), VE (-24,88 vs. -23,94%, p = 0,4150) sau VE/VO2 (-15,4 vs. -22,55%, p = 0,4643).
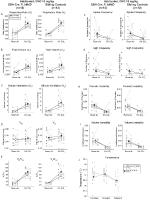
Figura 5. Ambele habituate DBH-Cre; RC::Animalele p_hm4d și grupurile de control frate administrate 10 mg/kg de CNO prezintă un deficit hipercapnic. După administrarea CNO, animalele DBH-Cre; RC::p_hm4d și controalele fraților nu prezintă nicio modificare a ventilației aerului din cameră și o reducere a RR (A), VE (C) și ve/VO2 (e) în condiții hipercapnice, fără modificări semnificative ale VT (B), VO2 (D), frecvența apneei (F), frecvența suspinului (G), instabilitatea periodică (H) sau a volumului (I) și temperatura (J).
animalele de tip sălbatic obișnuite și Neabituate injectate cu soluție salină nu au prezentat modificări Presalină și Postsalină
deși nu s-au observat fenotipuri la controalele injectate cu 1 mg/kg de CNO, am abordat posibilitatea ca injecția în sine să determine fenotipul observat la controalele injectate cu 10 mg/kg de CNO prin testarea animalelor de tip sălbatic obișnuite și neabituate injectate cu soluție salină (Figura 6). Atât în cohortele obișnuite, cât și în cele neobișnuite, animalele nu au prezentat nicio diferență în parametrii respiratori administrarea de presalină și postsalină. Nu s-au observat modificări semnificative ale frecvenței apneei, frecvenței suspinului, instabilității periodice sau a volumului sau temperaturii.
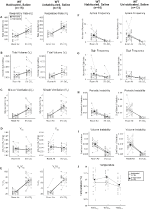
Figura 6. Atât animalele de tip sălbatic, cât și cele obișnuite, administrate cu soluție salină, nu prezintă nicio diferență între valorile pre – și post-injectare. După administrarea de soluție salină, atât animalele de tip sălbatic, cât și cele obișnuite nu au prezentat modificări în condițiile aerului din cameră sau hipercapnic în RR (A), VT (B), VE (C), VO2 (D), VE/VO2 (E), frecvența apneei (F), frecvența suspinului (G), instabilitatea periodică (H) sau a volumului (I) și temperatura (J).
raporturile concentrației clozapinei la CNO sunt mai mari în creier decât în ser
pentru a determina biodisponibilitatea CNO și clozapinei, am măsurat concentrațiile lor în ser și creier prin spectrometrie de masă. La treizeci de minute după o injecție intraperitoneală de CNO la șoareci, CNO se găsește într-o abundență relativă mai mică comparativ cu metabolitul său din spate clozapină în ser și creier pentru toate dozele testate, 0,1 mg/kg (ser p = 0,0054, creier p = 0,0001) (figura 7A), 1 mg/kg (ser p > 0,05, creier p = 0,0197) (figura 7B) și 10 mg/0.0036, creier p = 0.0005) (figura 7C). Când au fost analizate raporturile globale de clozapină la CNO, acestea au fost întotdeauna peste zero și au fost mai mari în creier decât în ser (vehicul p > 0,5, 0,1 mg/kg p > 0,5, 1 mg/kg p = 0,0018, 10 mg/kg p = 0,0160) (figura 7D).
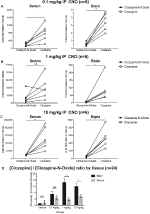
Figura 7. Concentrațiile de clozapină la CNO sunt mai mari în creier decât în ser. Probele de ser și creier au fost colectate de la șoareci de tip sălbatic la 30 de minute după administrarea CNO la 3 doze diferite de 0,1 mg/kg (a), 1 mg/kg (B), 10 mg/kg (C). Punctele de date individuale reprezintă abundența relativă de clozapină-N-oxid sau clozapină obținută de LC-MS.raportul clozapină-CNO pentru fiecare doză și SEM sunt, de asemenea, prezentate (D).
discuție
scopul inițial al acestui studiu a fost de a examina rolul neuronilor DBH-Cre în fiziologia respiratorie după perturbarea acută la animalele adulte neîngrădite și conștiente. Deoarece neuronii noradrenalinei și NA joacă un rol bine documentat în comportamentele de stres (Valentino și Van Bockstaele, 2008; Chen și colab., 2018), am căutat, de asemenea, să examinăm dacă obișnuința cu un protocol fiziologic stresant probabil ar avea un efect asupra fenotipurilor respiratorii observate anterior. În protocolul DREADD al pletismografiei întregului corp utilizat în laboratorul nostru și în altele, animalele sunt manipulate, sondate rectal pentru temperatură, expuse unui mediu nou (camera de pletismografie) și injectate intraperitoneal. Studiile anterioare au arătat că manipularea și obișnuința la injecție și alte proceduri „de rutină” pot modifica parametrii comportamentali și fiziologici, inclusiv respirația (Misslin și colab., 1982; Andrews și File, 1993; Lapin, 1995; Ryabinin și colab., 1999). Alți factori de stres aplicați modifică, de asemenea, respirația atât în condițiile de bază, cât și în cele de ventilație hipercapnică (Isom și Elshowihy, 1982; Kinkead și colab., 2001).
în studiile noastre care utilizează doze mari de CNO, am constatat că inhibarea mediată de hM4D a neuronilor NA definiți de DBH-Cre a dus la un reflex hipercapnic redus în VE/VO2 în patru cohorte experimentale definite, cu RR, VT și VE reduse, susținând studiile anterioare (Biancardi și colab., 2008; Viemari, 2008; Gargaglioni și colab., 2010). Cu toate acestea, subiecții de control cu frate obișnuit, cărora li s-a administrat o doză de CNO de 10 mg/kg, utilizată în studii respiratorii anterioare, au arătat un deficit de ventilație în condiții hipercapnice de aceeași magnitudine cu cea observată la animalele DBH-Cre; RC::P_hM4D. Niciun alt grup de control frate nu a prezentat acest fenotip, inclusiv cohorta obișnuită care a primit doar (1 mg/kg CNO) sau soluție salină. Aceste rezultate sugerează că dozele mai mari de CNO (10 mg/kg) au un efect asupra răspunsului hipercapnic care este demascat după o obișnuință extinsă, reducând în același timp nivelurile de stres la animale și că dozele mai mici de CNO nu au un efect asupra controlului respirator la animalele obișnuite. Aceste rezultate sunt, de asemenea, în acord cu munca de la Korsak și colab. OMS a demonstrat că doza mică de CNO (2 mg/kg) nu produce efecte în afara țintei în capacitatea de muncă într-un test de exerciții care a inclus antrenament anterior (Korsak și colab., 2018) și Fleury Curado și colab. OMS nu a prezentat efecte specifice CNO în doză mică (1 mg/kg) asupra debitului respirator (Fleury Curado și colab., 2018).
raporturile crescute ale nivelurilor de clozapină la CNO în ser și în creier (Figura 7) sunt în concordanță cu studiile recente care sugerează că CNO este ușor metabolizat înapoi la clozapină și prezintă o permeabilitate mai mare a creierului în comparație cu CNO la șoareci și în alte părți (Jann și colab., 1994; Chang și colab., 1998; Guettier și colab., 2009; Gomez și colab., 2017; Raper și colab., 2017). Cu toate acestea, nu este clar dacă efectele observate în afara țintei se datorează CNO sau clozapinei. Deoarece măsurătorile noastre hipercapnice au avut loc < 30 min după aplicarea CNO, este probabil ca efectele respiratorii în afara țintei să fie mediate de clozapină. Rezultatele noastre (Figura 7) arată niveluri relative ridicate de clozapină în creier, deși CNO nu este complet absent. Cu toate acestea, Huckstepp și colegii săi au folosit aplicația CNO directă la medulla ventrală la șobolanii anesteziați pentru a demonstra că numai la aerul din cameră și nu sub provocări hipercapnice sau hipoxice, aplicația CNO are un efect mic, crescând frecvența și scăzând durata expiratorie, dar lăsând VT neschimbată, fără efect clar observat în timpul hipercapniei (Huckstepp și colab., 2015). Având în vedere aplicarea directă la șobolani anesteziați și intervalul de timp al experimentelor, este probabil ca efectele mici în afara țintei observate să fie mediate de CNO și nu de clozapină.
metabolitul din spate clozapina este un medicament sedativ și antipsihotic utilizat în mod obișnuit în schizofrenie, cu multe ținte endogene, inclusiv acțiuni antagoniste cu afinitate scăzută la receptorii dopaminergici D1, D2 și D4, 5-HT2A, 5-HT2C, 5-HT3, 5-HT6 și 5-HT7, receptori serotoninergici H1 histaminici și receptori adrenergici de la 1996). Efectele în afara țintei observate aici pot rezulta din mecanisme și ținte distincte sau combinate. Clozapina poate afecta respirația ca sedativ. Un studiu anterior a arătat reducerea RR și VT Sub 5 și 10% CO2 atât în timpul somnului cu unde lente, cât și al stărilor de somn cu mișcare rapidă a ochilor, comparativ cu starea de veghe liniștită la șoareci (Nakamura și colab., 2007). Alternativ, inhibarea neuronilor vizați care exprimă DREADD poate duce la un efect antianxiogen sau anxiolitic similar cu protocolul nostru de obișnuință pentru a dezvălui efectele chemosenzoriale off-țintă mediate de CNO/clozapină. Ambele explicații sunt susținute de mai multe studii care au arătat că CO2 joacă un rol în răspunsurile de frică înnăscute și învățate și comportamentele legate de anxietate (Ziemann și colab., 2009; Feinstein și colab., 2013; Taugher și colab., 2014; Dlouhy și colab., 2015; Iarna și colab., 2017). Astfel, neuronii vizați în unele dintre aceste studii pot juca într-adevăr un rol în conducerea răspunsurilor comportamentale anxiogene, mai degrabă decât a reflexelor fiziologice chemosenzoriale, deoarece atât sistemele catecolaminergice, cât și cele serotoninergice sunt implicate în comportamentele de frică/anxietate și homeostazia chemosenzorială (Brust și colab., 2014; Hennessy și colab., 2017).
în schimb, fenotipurile chemosenzoriale observate cu niveluri ridicate de CNO pot fi autentice, deoarece am reușit să recapitulăm deficitul hipercapnic mediat de NA la doze CNO, o magnitudine de ordin mai mică la șoarecii obișnuiți, în timp ce grupurile de control nu au prezentat niciun efect CNO. Cu toate acestea, comparațiile complete între studiile anterioare sunt dificile din cauza lipsei, în unele cazuri, a datelor raportate VO2, VT, RR și VE/VO2. De exemplu, modificările temperaturii corpului sau ale ratei metabolice pot, de asemenea, să afecteze în mai multe moduri ieșirea respiratorie și chemosenzorială și că temperaturile camerei pletismografului au fost foarte diferite în unele cazuri (34 vs.30 centimetric C în studiile noastre), afectând intervalul dinamic al componentei barometrice a formei de undă și, prin urmare, volumul mareelor. În special, nu am văzut nici modificări apreciabile ale caracteristicilor formei de undă respiratorie în niciuna dintre condițiile noastre, nici stop cardio-respirator acut care a fost observat în studiile noastre anterioare cu doză mare, cu rombomeri întregi (Sun și colab., 2017).
rezultatele noastre arată pentru prima dată că CNO are un efect neașteptat asupra reflexului chemosenzorial hipercapnic care este demascat de o obișnuință extinsă. Important, în ciuda nivelurilor ridicate, un efect CNO în afara țintei a fost exclus anterior din cauza lipsei fenotipului în controalele fraților, dar pe care le arătăm devine clar după obișnuință. Oferim o caracterizare în afara țintei a CNO în sistemul model de șoarece pentru a completa studiile la șobolani și primate neumane. Rezultatele de aici ridică posibilitatea ca efecte suplimentare mediate de CNO, în afara țintei, asupra circuitelor studiate sau asupra circuitelor autonome sau homeostatice să existe, dar pot fi mascate în alte experimente controlate. Important, aceste date dezvăluie că anchetatorii ar trebui să se străduiască să utilizeze dozele minime de ligand activator posibil în combinație cu niveluri ridicate de obișnuință și că controalele adecvate trebuie incluse în manipulările genetice chimice pentru a aprecia și interpreta pe deplin datele experimentale.
materiale și metode
aprobare etică
studiile au fost aprobate de Colegiul de Medicină Baylor Comitetul instituțional pentru îngrijirea și utilizarea animalelor în conformitate cu protocolul AN-6171.
reproducerea, fondul Genetic și întreținerea șoarecilor
am menținut colonii ale tuturor tulpinilor noastre heterozigote de șoarece prin încrucișarea înapoi la șoareci c57bl/6j și tulpini homozigote prin cruci de frați. Pentru experimentele histologice, DBH-Cremice au fost împerecheate cu șoarecele homozigot AI9 (Madisen și colab., 2010) (JAX 007909). Pentru experimentele de pletismografie, șoarecii DBH-Cre au fost împerecheați cu homozigot RC::P_hM4D (Ray și colab., 2011) șoareci pentru a obține animale, în care toți șoarecii purtau alela RC::P_hM4D. Animalele frate care nu au moștenit alela Cre au fost folosite ca animale de control pentru descendenții cre pozitivi. Grundurile specifice Rosa26 pentru șoarecii AI9, RC::P_hM4D și RC::ePe au fost 5′-GCACTTGCTCTCCCAAAGTC, 5′-GGGCGTACTTGGCATATGAT și 5′-CTTTAAGCCTGCCAGAAGA și produc o bandă de 495 bp (vizată) și o bandă de 330 bp (wt). Primerii specifici Cre pentru toți șoferii cre rombomeri au fost 5 ‘- ATCGCCATCTTCCAGCAGGCGCACCATTGCCC și 5 ‘ – GCATTTCTGGGGATTGCTTA și au dat o bandă de 550 bp dacă este pozitivă. Pentru experimentele LC-MS, șoarecii c57bl/6j au fost obținuți de la Centrul de Medicină comparată (CCM), Colegiul de Medicină Baylor.
Histologie
DBH-Cre cu vârsta cuprinsă între patru și opt săptămâni; șoarecii adulți Ai9 au fost perfuzați transcardial cu soluție salină tamponată cu fosfat de 0,1 m (PBS) și apoi cu paraformaldehidă 4% (PFA) în PBS. Creierele au fost disecate și fixate pentru 2 ore în 4% PFA înainte de o clătire PBS și echilibrare în 20% zaharoză în PBS. Creierele au fost apoi secționate în secțiuni de 30 de centimetri, montate pe diapozitive și etichetate cu anticorpi imunofluorescenți. Am colorat peste noapte cu anticorp anti-tirozină hidroxilază pentru a identifica neuronii catecolaminergici (1:1.000, Millipore AB152) urmat de 2 ore cu măgar anti-iepure Cy3 secundar (1:500, Jackson 711-165-152) în 0,1% Triton-X în PBS (PBST) cu 5% ser de măgar. Imaginile au fost colectate pe un microscop Epifluorescent vertical Zeiss.
electrofiziologie
feliile orizontale de creier care conțineau locus coeruleus (300 unqq) au fost tăiate cu un vibratom (Leica VT 1000s, Leica Microsystems, Buffalo Grove, IL) de la șoareci adulți DBH-Cre; RR2P; RC::Epe (~1 lună) în 4 LCR (ACSF). Feliile au fost scufundate într-o cameră perfuzată cu ACSF oxigenat (95% O2 și 5% CO2) conținând în mM: 124 Naci, 2,0 KCl, 1,3 MgSO4, 2,5 CaCl2, 1,2 KH2PO4, 25 NaHCO3 și 10 glucoză (2-3 ml/min). Înregistrările cu celule întregi au fost efectuate la 30 de Centimetre C folosind tehnici convenționale de fixare a plasturelui și un amplificator MultiClamp 700B (Molecular Devices, Union City, CA). Neuronii pozitivi GFP din locus coeruleus au fost identificați vizual și ulterior fotografiați prin video de contrast cu interferență diferențială în infraroșu pe scena unui microscop vertical (Axioskope FS2, Carl Zeiss, Oberkochen, Germania). Pipetele de plasture (rezistențe 4-6 MU) au fost umplute cu (în mM): 110 K-gluconat, 10 KCI, 10 HEPES, 10 NA2-fosfocreatină, 2 Mg3-ATP și 0,2 Na3-GTP; pH-ul a fost ajustat la 7,2 și osmolaritatea la 300 mOsm. Potențialul de deținere a fost de -70 mV. CNO a fost aplicat baie.
pletismografie
pletismografie pe șoareci conștienți și neîngrădiți a fost efectuată așa cum este descris pe animale adulte cu vârsta cuprinsă între 6 și 12 săptămâni (Ray și colab., 2011). Șoarecii obișnuiți au fost supuși unui protocol de obișnuință de 5 zile, fiecare zi constând în câteva minute de manipulare, temperatură luată de sonda rectală, injecție salină intraperitoneală și 30 de minute în camera de pletismografie. Șoarecii au fost apoi testați în decurs de 1 săptămână de la ultima zi de obișnuință. Șoarecii nevătămați nu au fost expuși la manipulare sau la camera de pletismografie. Toți șoarecii au fost na la CNO și au fost utilizați o singură dată.
în ziua testării, șoarecii au fost luați din cușca lor de origine, cântăriți și temperatura rectală a fost luată. Animalele au fost apoi plasate într-o cameră de pletismografie cu debit, cu temperatură controlată (cu manta de apă la 30 C) și lăsate să se aclimatizeze timp de cel puțin 20 de minute în condiții de aer din cameră (21% O2/79% N2). După aclimatizare și măsurare sub aerul din încăpere, gazul din cameră a fost trecut la un amestec hipercapnic de 5% CO2/21% O2/74% N2 timp de 20 min. Gazul camerei a fost apoi comutat înapoi în aerul camerei timp de 20 de minute. Șoarecii au fost îndepărtați pentru scurt timp pentru măsurarea temperaturii rectale și injectarea intra-peritoneală a CNO (Institutul Național de sinteză chimică a sănătății mintale și programul de aprovizionare cu medicamente) dizolvat în soluție salină (1 sau 0,1 mg/ml) pentru o concentrație eficientă de 10 sau 1 mg/kg, respectiv. Animalul a fost returnat în cameră pentru încă 20 de minute de aer din cameră, 20 de minute de hipercapnie și 20 de minute de aer din cameră. Animalul a fost apoi scos din cameră, iar temperatura rectală a fost luată imediat după terminarea experimentului.
cromatografie lichidă-spectrometrie de masă
douăzeci și patru de șoareci de tip sălbatic, împărțiți uniform pe sex, au fost cântăriți și tratați cu 10 mg/kg CNO, 1 mg/kg CNO, 0,1 mg/kg CNO sau vehicul. La treizeci de minute după injectare, probele de sânge au fost colectate prin puncție cardiacă și plasate în Microtainere BD. Probele au fost centrifugate la 4 C la 13.500 rpm în centrifugă de vârf și s-au colectat supernatanți. Serul și creierul au fost păstrate la -20 centimetric C până la extracție.
solvenți de calitate HPLC apă, acetonitril cloroform și metanol și acid formic de calitate spectrometrică de masă au fost obținute de la Sigma-Aldrich (St.Louis, MO). Soluția de calibrare care conține mai mulți calibranți într-o soluție de acetonitril, acid trifluoracetic și apă a fost achiziționată de la Agilent Technologies (Santa Cruz, CA). Metaboliții și standardele interne, inclusiv acidul N-acetil aspartic-d3, triptofanul-15n2, acidul glutamic-d5, timina-d4, acidul gibberelic, trans-zeatina, acidul jasmonic, acidul antranilic 15N și testosteronul-D3 au fost achiziționate de la Sigma-Aldrich (St.Louis, MO). Microtainer R SST TM a fost obținut de la Becton Dickinson (Franklin Lakes, NJ).
extracția a constat din 750 unkticli de metanol rece ca gheața:apă (4:1) conținând 20 de standarde interne cu vârf de unkticli care au fost adăugate la fiecare probă de creier (50 mg) și controale de calitate și apoi au fost omogenizate timp de 1 min fiecare. Apoi, 750 unktil de 100% acetonitril conținând 20 unktil standarde interne cu vârf au fost adăugate la proba de spălare (100 unktill) și controale de calitate și apoi sonicat timp de 5 min. Toate probele au fost centrifugate la 5.000 rpm Timp de 10 min la 4 C. Supernatantul rezultat a fost colectat și au fost injectate 20 unqql în LC-MS.
toate probele au fost analizate folosind spectrometru de masă triplu quadrupol 6,490 (Agilent Technologies, Santa Clara, CA) cuplat la sistemul HPLC (Agilent Technologies, Santa Clara, CA) prin monitorizarea reacțiilor multiple (MRM). Au fost obținute aproximativ 8-11 puncte de date pentru fiecare metabolit detectat. Metaboliții detectați au fost clozapina, CNO și norclozapina (N-desmetil clozapina). Modul pozitiv ESI a fost utilizat în metodă. Coloana HPLC a fost Coloana ACQUITYUPLC C18 (100, 1,8, 1,8 și 2,1 mm, 100 mm. Milford, MA, SUA) cu un debit de 0,5 ml/min.
colectarea și analiza datelor
pletismografie
pletismografie schimbările de presiune au fost măsurate folosind un traductor de presiune diferențială Validyne DP45 și o cameră de referință și un demodulator purtător CD15 și înregistrate cu LabChartPro în timp real. Formele de undă au fost analizate offline folosind LabChartPro și cod MATLAB scris personalizat pentru a determina rata respiratorie (RR), volumul mareelor (VT) (Ray și colab., 2011), ventilație minute (VE) și analiza modelului. Formele de undă respiratorii au fost colectate offline în perioadele în care animalul era în repaus, iar citirile erau libere de artefacte de mișcare. Au fost analizate un minim de 1 min date cumulative compilate din urme de cel puțin 10 s Lungime din ultimele 10 min ale unei condiții experimentale date. Consumul de O2 a fost determinat prin compararea compoziției gazului între calibrarea într-o cameră goală și respirația vie folosind un senzor de oxigen și analizor aei. Temperatura camerei a fost monitorizată constant cu ajutorul unui ThermoWorks MicroThermo 2 și sondă și a fost înregistrată cu LabChartPro în timp real.
Poincar masuratorile si frecventa de suspin si apnee au fost determinate folosind 1 min de urme fara miscare din fiecare stare de respiratie. Suspinele au fost definite ca o respirație cu amplitudine de cel puțin două ori mai mare decât respirația medie. Apneele au fost definite ca un interval interbreath (IBI) cel puțin de două ori mai mare decât media IBI. Coeficientul de variație (CV) al IBI și amplitudine a fost, de asemenea, calculat din aceeași compilație de urme de 1-min a fiecărei condiții de respirație (eroare standard IBI sau amplitudine/medie IBI sau amplitudine).
statistici
pletismografie
rezultatele (RR, VT, VE, VO2, VE/VO2, numărul de apnee și suspine și CVs de IBI și amplitudine) pentru aerul din cameră și datele hipercapnice au fost comparate între cohortele DBH-Cre; RC::P_hM4D și controalele frate folosind un model liniar de regresie cu efecte mixte cu tipul animal (experimental vs.control) și administrarea CNO (pre vs. post injectare) ca efecte fixe și ID animal ca efect aleatoriu. Datele de temperatură au fost comparate folosind un model liniar de regresie cu efecte mixte cu tipul animal (experimental vs.control) ca efect fix. Un p < 0.05 a fost utilizat pentru a indica semnificația statistică, iar punctele de date individuale, media și eroarea standard a mediei sunt afișate pe toate diagramele.
cromatografie lichidă-spectrometrie de masă
aria obținută sub vârf pentru fiecare probă a fost normalizată prin control intern și apoi la vehicul înainte de efectuarea analizei statistice. Testul t nepereche a fost utilizat pentru a compara abundența relativă a clozapinei și CNO în fiecare țesut în funcție de grupul de concentrație.
declarație de etică
toate experimentele au fost aprobate de Comitetul instituțional pentru îngrijirea și utilizarea animalelor al Colegiului de Medicină Baylor. Experimentele s-au conformat Standardelor Naționale pentru îngrijirea și utilizarea animalelor experimentale stabilite de Asociația pentru evaluarea și acreditarea îngrijirii animalelor de laborator.
contribuțiile autorului
JS, FS-M, MC-M și RR au conceput și proiectat experimentele. JS, FS-M și PZ au efectuat experimentele și au contribuit la analiza datelor. JS, FS-M, VM, MC-M și RR au scris lucrarea.
finanțare
acest studiu a fost susținut de Granturile R01HL130249 și R01HL130249-S1 de la Institutul Național al Inimii, Plămânului și sângelui; Premiul March of Dimes Basil O ‘ Connor; și Institutul Medical McNair.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
mulțumim Drs.Shaila K. Mani și Benjamin Arenkiel pentru discuții utile. Mulțumim, de asemenea, Colegiului de Medicină Baylor Proteomics core facility pentru executarea cromatografiei lichide-spectrometrie de masă.
Amiel, J., Dubreuil, V., Ramanantsoa, N., Fortin, G., Gallego, J., Brunet, J.-F. și colab. (2009). PHOX2B în controlul respirator: lecții din sindromul hipoventilației centrale congenitale și modelele sale de șoarece. Respir. Physiol. Neurobiol. 168, 125–132. doi: 10.1016 / j.resp.2009.03.005
CrossRef text integral / Google Scholar
Andrews, N. și File ,S. E. (1993). Istoricul manipulării șobolanilor modifică efectele comportamentale ale medicamentelor în testul ridicat plus-labirint al anxietății. Eur. J. Pharmacol. 235, 109–112. doi: 10.1016/0014-2999(93)90827-5
PubMed rezumat / CrossRef Text Complet / Google Scholar
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S. și Roth, B. L. (2007). Evoluând blocarea pentru a se potrivi cheii pentru a crea o familie de receptori cuplați cu proteina G puternic activați de un ligand inert. Proc. Natl. Acad. Sci. SUA 104, 5163-5168. doi: 10.1073 / pnas.0700293104
PubMed rezumat / CrossRef text integral / Google Scholar
Ashby, C. R. și Wang, R. Y. (1996). Acțiuni farmacologice ale medicamentului antipsihotic atipic clozapină: o revizuire. Sinapsa 24, 349-394. doi: 10.1002 / (SICI)1098-2396(199612)24:4<349::AID-SYN5>3.0.CO; 2-D
rezumat PubMed / CrossRef Text Complet / Google Scholar
Bahia, C. M. C. d.S. și Pereira, J. S. (2015). Apneea obstructivă de somn și bolile neurodegenerative: o relație bidirecțională. Dement. Neuropsihol. 9, 9–15. doi: 10.1590 / S1980-57642015DN91000003
PubMed rezumat / CrossRef text integral / Google Scholar
Biancardi, V., B, K. C., Almeida, M. C., și Gargaglioni, L. H. (2008). Neuronii noradrenergici locus coeruleus și CO2 conduc la respirație. Pflugers Arch. – Eur. J. Physiol. 455, 1119–1128. doi: 10.1007 / s00424-007-0338-8
PubMed rezumat / CrossRef Text Complet / Google Scholar
Brust, R. D., Corcoran, A. E., Richerson, G. B., Nattie, E. și Dymecki, S. M. (2014). Identificarea funcțională și de dezvoltare a unui subtip molecular al neuronului serotoninergic cerebral specializat în reglarea dinamicii respirației. Celula Rep. 9, 2152-2165. doi: 10.1016 / j. celrep.2014.11.027
PubMed rezumat / CrossRef text integral / Google Scholar
Chang, W. H., Lin, S. K., Lane, H. Y., Wei, F. C., Hu, W. H., Lam, Y. W. și colab. (1998). Metabolizarea reversibilă a clozapinei și a n-oxidului de clozapină la pacienții cu schizofrenie. Prog. Neuro-Psihofarmacol. Biol. Psihiatrie 22, 723-739. doi: 10.1016 / S0278-5846(98)00035-9
PubMed rezumat / CrossRef Text Complet / Google Scholar
Cheng, Q., Plummer, N. W., Smith, K. G., și colab. (2018). Identificarea genetică a unei populații de neuroni noradrenergici implicați în atenuarea răspunsurilor legate de stres. Mol. Psihiatrie 1-16. doi: 10.1038 / s41380-018-0245-8
PubMed rezumat / CrossRef Text Complet / Google Scholar
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kawasaki, H., Oya, H., Buzza, C. și colab. (2015). Respirația inhibată atunci când convulsiile se răspândesc în amigdala și la stimularea amigdalei. J. Neurosci. 35, 10281–10289. doi: 10.1523/JNEUROSCI.0888-15. 2015
PubMed rezumat / CrossRef text integral / Google Scholar
Feinstein, J. S., Buzza, C., Hurlemann, R., Follmer, R. L., Dahdaleh, N. S., Coryell, W. H. și colab. (2013). Frica și panica la om cu leziuni amigdale bilaterale. Nat. Neuroști. 16, 270–272. doi: 10.1038 / nn.3323
rezumat PubMed / CrossRef Text Complet / Google Scholar
Fitton, A. și Heel, R. C. (1990). Clozapină. O revizuire a proprietăților sale farmacologice și a utilizării terapeutice în schizofrenie. Droguri 40, 722-747. doi: 10.2165/00003495-199040050-00007
PubMed rezumat / CrossRef Text Complet / Google Scholar
Fleury Curado, T. A., Pho, H., Dergacheva, O., Berger, S., Lee, R., Freire, C. și colab. (2018). Tăcerea motoneuronilor hipoglosali duce la respirația dezordonată a somnului la șoarecii slabi. În față. Neurol. 9:962. doi: 10.3389 / fneur.2018.00962
CrossRef Text Complet / Google Scholar
Gargaglioni, L. H., Hartzler, L. K., și Putnam, R. W. (2010). Locus coeruleus și chemosensibilitate centrală. Respir. Physiol. Neurobiol. 173, 264–273. doi: 10.1016 / j.resp.2010.04.024
PubMed rezumat / CrossRef text integral / Google Scholar
Gomez, J. L., Bonaventura, J., Lesniak, W., Mathews, W. B., Sysa-Shah, P., Rodriguez, L. A. și colab. (2017). Chemogenetica a dezvăluit: ocuparea DREADD și activarea prin clozapină convertită. Știință 357, 503-507. doi: 10.1126 / știință.aan2475
rezumat PubMed / CrossRef Text Complet / Google Scholar
Guettier, J. M., Gautam, D., Scarselli, M., Ruiz de Azua, I., Li, J. H., Rosemond, E. și colab. (2009). O abordare Chimico-genetică pentru a studia reglarea proteinei g a funcției celulelor beta in vivo. Proc. Natl. Acad. Sci. SUA 106, 19197-19202. doi: 10.1073 / pnas.0906593106
CrossRef Text Complet / Google Scholar
Hakim, F., Gozal, D. și Kheirandish-Gozal, L. (2012). Modificări simpatice și catecolaminergice în apneea de somn, cu accent deosebit pe copii. În față. Neurol. 3:7. doi: 10.3389 / fneur.2012.00007
rezumat PubMed / CrossRef Text Complet / Google Scholar
Hennessy, M. L., Corcoran, A. E., Brust, R. D., Chang, Y., Nattie, E. E. și Dymecki, S. M. (2017). Activitatea neuronilor raphe Pet1 care exprimă Tahikinină1 modulează chemoreflexul respirator. J. Neurosci. 37, 1807–1819. doi: 10.1523/JNEUROSCI.2316-16. 2016
rezumat PubMed / CrossRef Text Complet / Google Scholar
Huckstepp, R. T. R., Cardoza, K. P., Henderson, L. E. și Feldman, J. L. (2015). Rolul nucleilor parafaciali în controlul respirației la șobolanii adulți. J. Neurosci. 35, 1052–1067. doi: 10.1523/JNEUROSCI.2953-14. 2015
rezumat PubMed / CrossRef Text Complet / Google Scholar
Ilg, A.-K., Enkel, T., Bartsch, D. și B Inkthner, F. (2018). Efectele comportamentale ale clozapinei sistemice acute cu doze mici la șobolanii de tip sălbatic: implicații pentru utilizarea Dreadd-urilor în neuroștiința comportamentală. În față. Comportă-te. Neuroști. 12:173. doi: 10.3389 / fnbeh.2018.00173
CrossRef Text Complet / Google Scholar
Isom, G. E. și Elshowihy, R. M. (1982). Interacțiunea stresului acut și cronic cu respirația: modificarea prin naloxonă. Pharmacol. Biochem. Comportă-te. 16, 599–603. doi: 10.1016/0091-3057(82)90422-1
PubMed rezumat / CrossRef Text Complet / Google Scholar
Jann, M. W., Lam, Y. W. și Chang, W. H. (1994). Formarea rapidă a clozapinei la cobai și om după administrarea de clozapină-N-oxid. Arch. Int. Pharmacodyn. Acolo. 328, 243–250.
Rezumat PubMed / Google Scholar
Joober, R. (2010). Clozapină: o moleculă distinctă, puțin înțeleasă și subutilizată. J. Neurologie Psihiatrică. 35, 147–149. doi: 10.1503/jpn.100055
rezumat PubMed / CrossRef Text Complet / Google Scholar
Kinkead, R., Dupenloup, L., Valois, N. și Gulemetova, R. (2001). Atenuarea indusă de stres a răspunsului ventilator hipercapnic la șobolanii treji. J. Appl. Physiol. 90, 1729–1735. doi: 10.1152 / jappl.2001.90.5.1729
PubMed Rezumat / CrossRef Text Integral / Google Scholar
Klein, D. F. (1993). Alarme false de sufocare, panică spontană și condiții conexe. O ipoteză Integrativă. Arch. Gen. Psihiatrie 50, 306-317. doi: 10.1001/archpsyc.1993.01820160076009
PubMed rezumat / CrossRef text integral / Google Scholar
Korsak, A., Sheikhbahaei, S., Machhada, A., Gourine, A. V. și Huckstepp, R. T. R. (2018). Rolul neuronilor parafaciali în controlul respirației în timpul exercițiilor fizice. Sci. Rep. 8:400. doi: 10.1038 / s41598-017-17412-z
CrossRef Text Complet / Google Scholar
Lapin, I. P. (1995). Numai controale: efectul manipulării, injecției false și injecției intraperitoneale de soluție salină asupra comportamentului șoarecilor într-un labirint plus ridicat. J. Pharmacol. Toxicol. Metode 34, 73-77. doi: 10.1016 / 1056-8719 (95)00025-D
PubMed rezumat / CrossRef textul integral / Google Scholar
Lavezzi, A. M., Alfonsi, G. și Matturri, L. (2013). Fiziopatologia complexului locus coeruleus uman în moartea subită inexplicabilă fetală/neonatală. Neurol. Rezoluția 35, 44-53. doi: 10.1179 / 1743132812Y. 0000000108
PubMed rezumat / CrossRef text integral / Google Scholar
MacLaren, D. A. A., Browne, R. W., Shaw, J. K., Krishnan Radhakrishnan, S., Khare, P., Espa Xusta, R. A. și colab. (2016). Administrarea de N-oxid de clozapină produce efecte comportamentale la șobolanii long-evans: implicații pentru proiectarea experimentelor DREADD. eNeuro 3, 1-14. doi: 10.1523 / ENEURO.0219-16. 2016
PubMed rezumat / CrossRef text integral / Google Scholar
Madisen, L., Zwingman, T. A., Sunkin, S. M., Oh, S. W., Zariwala, H. A., Gu, H. și colab. (2010). Un sistem de raportare și caracterizare cre robust și cu randament ridicat pentru întregul creier al mouse-ului. Nat. Neuroști. 13, 133–140. doi: 10.1038 / nn.2467
rezumat PubMed / CrossRef Text Complet / Google Scholar
Mahler, S. V. și Aston-Jones, G. (2018). CNO evil? Considerații pentru utilizarea dreadds în neuroștiința comportamentală. Neuropsihofarmacologie 43, 934-936. doi: 10.1038 / npp.2017.299
rezumat PubMed / CrossRef Text Complet / Google Scholar
manvich, D. F., Webster, K. A., Foster, S. L., Farrell, ms, Ritchie, J. C., Porter, J. H. și colab. (2018). DREADD agonist clozapină N-oxid (CNO) este metabolizat invers la clozapină și produce efecte de stimulare interoceptivă asemănătoare clozapinei la șobolani și șoareci. Sci. Rep. 8, 3840. doi: 10.1038 / s41598-018-22116-z
CrossRef Text Complet / Google Scholar
Misslin, R., Herzog, F., Koch, B. și Ropartz, P. (1982). Efectele izolării, manipulării și noutății asupra hipofizei — răspunsul suprarenal la șoarece. Psihoneuroendocrinologie 7, 217-221. doi: 10.1016/0306-4530(82)90015-4
PubMed rezumat / CrossRef Text Complet / Google Scholar
Nakamura, A., Zhang, W., Yanagisawa, M., Fukuda, Y. și Kuwaki, T. (2007). Vigilență atenuarea dependentă de stat a hipercapnic chemoreflex și apneea de somn exagerată la șoarecii knockout orexin. J. Appl. Physiol. 102, 241–248. doi: 10.1152 / japplphysiol.00679.2006
PubMed rezumat / CrossRef text integral / Google Scholar
Obonai, T., Yasuhara, M., Nakamura, T. și Takashima, S. (1998). Modificarea neuronilor catecolaminici în trunchiul cerebral al victimelor sindromului morții subite a sugarului. Pediatrie 101, 285-288. doi: 10.1542 / peds.101.2.285
rezumat PubMed / CrossRef Text Complet / Google Scholar
Ozawa, Y., Takashima, S. și Tada, H. (2003). Modificări ale subtipului receptorului alfa2-adrenergic în trunchiul cerebral în sindromul morții subite a sugarului. Zumzet Devreme. Dev. 75 (Suppl.), S129–S138. doi: 10.1016 / j. pathophys.2004.01.015
CrossRef text integral / Google Scholar
Padovan-Hernandez, Y. și Knackstedt, L. A. (2018). Reducerea dependentă de doză a locomoției induse de cocaină de clozapină-N-oxid la șobolani cu antecedente de autoadministrare a cocainei. Neuroști. Let. 674, 132–135. doi: 10.1016 / j. neulet.2018.03.045
rezumat PubMed / CrossRef Text Complet / Google Scholar
Ramanantsoa, N., Hirsch, M.-R., Thoby-Brisson, M., Dubreuil, V., Bouvier, J., Ruffault, P.-L. și colab. (2011). Respirație fără chemosensibilitate CO (2) la mutanții condiționali Phox2b. J. Neurosci. 31, 12880–12888. doi: 10.1523/JNEUROSCI.1721-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Raper, J., Morrison, R. D., Daniels, J. S., Howell, L., Bachevalier, J., Wichmann, T., et al. (2017). Metabolism and distribution of clozapine-N-oxide: implications for nonhuman primate chemogenetics. ACS Chem. Neurosci. 8, 1570–1576. doi: 10.1021/acschemneuro.7b00079
PubMed Abstract | CrossRef Full Text | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Soriano, L. P., Nattie, E. E., and Dymecki, S. M. (2013). Egr2-neuronii controlează răspunsul respirator adult la hipercapnie. Brain Res. 1511, 115-125. doi: 10.1016 / j. brainres.2012.12.017
PubMed rezumat / CrossRef text integral / Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Kim, J. C., Richerson, G. B., Nattie, E. și colab. (2011). Controlul respirator și al temperaturii corporale afectat la inhibarea acută a neuronilor serotoninergici. Știință 333, 637-642. doi: 10.1126 / știință.1205295
Rezumat PubMed / CrossRef Text Complet / Google Scholar
Ryabinin, A. E., Wang, Y. M., și Finn, D. A. (1999). Niveluri diferite de imunoreactivitate Fos după manipularea repetată și stresul injectabil la două tulpini consangvinizate de șoareci. Pharmacol. Biochem. Comportă-te. 63, 143–151. doi: 10.1016 / S0091-3057(98)00239-1
PubMed rezumat / CrossRef Text Complet / Google Scholar
Snyder, B., coajă, B., Cunningham, J. T. și Cunningham ,R. L. (2017). Hipoxia intermitentă cronică induce stresul oxidativ și inflamația în regiunile creierului asociate cu neurodegenerarea în stadiu incipient. Physiol. Rep. 5, 1-13. doi: 10.14814 / phy2.13258
rezumat PubMed / CrossRef Text Complet / Google Scholar
Sun, J. J., Huang, T.-W., Neul, J. L. și Ray, R. S. (2017). Genele de modelare a creierului posterior embrionar delimitează populații homeostatice cardio-respiratorii și metabolice distincte la adult. Sci. Rep. 7: 9117. doi: 10.1038 / s41598-017-08810-4
CrossRef Text Complet / Google Scholar
Sun, J. J. și Ray, R. S. (2017). Tg (Th-Cre)FI172Gsat (Th-Cre) definește neuronii care sunt necesari pentru reflexele hipercapnice și hipoxice complete. Biol. Deschis 6, 1200-1208. doi: 10.1242 / bio.026823
PubMed rezumat / CrossRef Text Complet / Google Scholar
Taugher, R. J., Lu, Y., Wang, Y., Kreple, C. J., Ghobbeh, A., Fan, R. și colab. (2014). Nucleul de pat al stria terminalis este esențial pentru comportamentul legat de anxietate evocat de CO2 și acidoză. J. Neurosci. 34, 10247–10255. doi: 10.1523/JNEUROSCI.1680-14. 2014
rezumat PubMed / CrossRef Text Complet / Google Scholar
Valentino, R. J. și Van Bockstaele, E. (2008). Reglarea convergentă a activității locus coeruleus ca răspuns adaptiv la stres. Eur. J. Pharmacol. 583, 194–203. doi: 10.1016 / j. ejfar.2007.11.062
CrossRef text integral / Google Scholar
Verbraecken, J. și McNicholas, W. T. (2013). Mecanica respiratorie și controlul ventilator în sindromul de suprapunere și hipoventilația obezității. Respir. Res. 14: 132. doi: 10.1186/1465-9921-14-132
PubMed rezumat / CrossRef Text Complet / Google Scholar
Viemari, J.-C. (2008). Modularea noradrenergică a rețelei neuronale respiratorii. Respir. Physiol. Neurobiol. 164, 123–130. doi: 10.1016 / j.resp.2008.06.016
rezumat PubMed / CrossRef Text Complet / Google Scholar
iarna, A., Ahlbrand, R., Naik, D. și Sah, R. (2017). Sensibilitate comportamentală diferențiată la inhalarea dioxidului de carbon (CO2) la șobolani. Neuroștiințe 346, 423-433. doi: 10.1016 / j. neuroștiințe.2017.01.003
PubMed rezumat / CrossRef text integral / Google Scholar
Ziemann, A. E., Allen, J. E., Dahdaleh, N. S., Drebot, I. I., Coryell, M. W., Wunsch, A. M. și colab. (2009). Amigdala este un chemosenzor care detectează dioxidul de carbon și acidoza pentru a provoca un comportament de frică. Cell 139, 1012–1021. doi: 10.1016/j.cell.2009.10.029
PubMed Abstract | CrossRef Full Text | Google Scholar