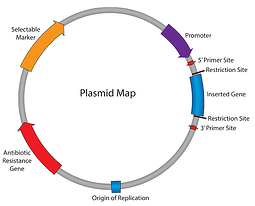
 do tej pory w naszej serii plazmidów 101 pracowaliśmy nad mapą plazmidów: opornością na antybiotyki, pochodzeniem replikacji i tak dalej. Do tego momentu możemy replikować nasz plazmid i upewnić się, że komórki go utrzymują; następnym krokiem jest uzyskanie plazmidu, aby wyrazić nasz interesujący Gen. Wprowadź promotor — element odpowiedzialny za inicjację transkrypcji Insertu do RNA.
do tej pory w naszej serii plazmidów 101 pracowaliśmy nad mapą plazmidów: opornością na antybiotyki, pochodzeniem replikacji i tak dalej. Do tego momentu możemy replikować nasz plazmid i upewnić się, że komórki go utrzymują; następnym krokiem jest uzyskanie plazmidu, aby wyrazić nasz interesujący Gen. Wprowadź promotor — element odpowiedzialny za inicjację transkrypcji Insertu do RNA.
w praktyce termin „promotor” opisuje połączenie promotora (miejsca wiązania polimerazy RNA) i operatorów (elementów odpowiedzi). Promotory mają długość około 100 do 1000 par zasad i znajdują się przed ich docelowymi genami. Sekwencja regionu promotora kontroluje Wiązanie polimerazy RNA i czynników transkrypcyjnych, dlatego promotory odgrywają dużą rolę w określaniu, gdzie i kiedy będzie wyrażany interesujący Gen.
polimeraza RNA
RNA jest transkrybowana z DNA przy użyciu polimerazy RNA (RNAP). U bakterii odbywa się to za pomocą jednego enzymu, jednak u eukariotów występują wielolipowe polimerazy, z których każda jest odpowiedzialna za określony podzbiór RNA. Aby uzyskać tę swoistość, EUKARIOTYCZNY RNAP może rozpoznawać i wiązać się z określonymi elementami promotora. Oznacza to, że promotor obecny w kręgosłupie plazmidu musi być zgodny z typem RNA, który należy wytworzyć: jeśli chcesz uzyskać mRNA (do ekspresji genu), musisz użyć promotora RNAP II, podczas gdy małe RNA (takie jak shRNA) są transkrybowane z promotorów RNAP III. Ten post zawiera promotory dla ogólnej transkrypcji RNAP II i RNAP III; jednak używanie wirusowych ltr jako promotorów RNAP II jest powszechnie stosowane w konstrukcjach lentiwirusowych i retrowirusowych i omówimy je w przyszłym poście na temat wirusowych części wektorowych.
swoistość promotora
oprócz wyboru promotora na podstawie rodzaju transkryptu RNA, należy również upewnić się, że plazmid ma promotor odpowiedni do działania w organizmie gospodarza. Ponieważ maszyny transkrypcyjne różnią się między typami komórek lub organizmami, promotory muszą być podobnie zmienne. Promotory bakteryjne działają tylko w komórkach prokariotycznych i zazwyczaj tylko u tych samych lub blisko spokrewnionych gatunków, z których pochodzą. Podobnie różne typy komórek eukariotycznych (Ssaki, drożdże, rośliny itp.) wymagają unikalnych promotorów i istnieje bardzo mało krzyżowania. Ogólnie rzecz biorąc, promotory u bakterii są mniej zróżnicowane i złożone, mają mniej części niż w komórkach eukariotycznych. Niektóre promotory są konstytutywnie aktywne i włączone przez cały czas, podczas gdy inne są dokładniej kontrolowane. Regulowane promotory mogą działać tylko w niektórych tkankach lub w określonych okresach rozwoju lub mogą istnieć sposoby włączania lub wyłączania ich do woli za pomocą substancji chemicznej, ciepła lub światła. W komórce same promotory są kontrolowane przez jeszcze inne czynniki regulacyjne: wzmacniacze, elementy graniczne, izolatory i tłumiki; jednak może wystąpić „wyciek” transkrypcji. Zwykle nie jest to duży problem dla komórek, ale może zakłócać wyniki badań lub nawet zabijać komórki, jeśli interesujący Cię gen jest toksyczny. Aby temu przeciwdziałać, naukowcy stworzyli syntetyczne promotory, które zazwyczaj zawierają pewną kombinację innych elementów promotora i są bardziej ściśle regulowane.
wspólne promotory dla eukariotów i prokariotów
poniżej zamieściliśmy dwie tabele referencyjne wymieniające niektóre z najczęstszych promotorów bakteryjnych i ssaków. Te listy nie są bynajmniej wyczerpujące,ale powinny być dobrym miejscem, aby zacząć od próby wybrania idealnego organizatora!
promotory eukariotyczne
| promotor | transkrypcja RNA | opis | ekspresja | Uwagi dodatkowe | |
| CMV | ekspresja Ogólna | mRNA | silny promotor ekspresji ssaków z ludzkiego wirusa cytomegalii | konstytutywny | może zawierać region wzmacniający. Może być wyciszony w niektórych typach komórek. |
| EF1a | ogólna ekspresja | mRNA | silna ekspresja ssaków z ludzkiego czynnika wydłużenia 1 alfa | konstytutywny | ma tendencję do uzyskania spójnej ekspresji niezależnie od rodzaju komórki lub fizjologii. |
| SV40 | ekspresja Ogólna | mRNA | promotor ekspresji ssaków z Małpiego wirusa wakuolizującego 40 | konstytutywny | może zawierać wzmacniacz. |
| PGK1 (u ludzi lub myszy) | ekspresja Ogólna | mRNA | promotor ssaków z genu kinazy fosfogliceratowej. | Ma tendencję do opierania się regulacji promotora w dół z powodu metylacji lub deacetylacji. | |
| Ubc | ekspresja Ogólna | mRNA | promotor ssaków z ludzkiego genu ubikwityny C | konstytutywny | jak sama nazwa wskazuje, promotor ten jest wszechobecny. |
| ludzka beta aktyna | ekspresja Ogólna | mRNA | promotor ssaków z genu beta aktyny | Wersja z kurczakiem jest powszechnie stosowana w Hybrydach promotorów. | |
| CAG | ekspresja Ogólna | mRNA | silny hybrydowy promotor ssaków | konstytutywny | zawiera wzmacniacz CMV, promotor beta-aktyny z kurczaka i akceptor splotu beta-globiny królika. |
| tre | ekspresja Ogólna | mRNA | promotor elementu odpowiedzi tetracykliny | indukowany Tetracyliną lub jej pochodnymi. | zazwyczaj zawiera minimalny promotor o małej aktywności podstawowej i kilka operatorów tetracyklin. Transkrypcja może być włączana lub wyłączana w zależności od tego, w jaki sposób tet transactivator jest używany. |
| UAS | ogólna ekspresja | mRNA | promotor Drosophila conaining gal4 binding sites | Specific | wymaga obecności genu Gal4 do aktywacji promotora. |
| AC5 | ogólna ekspresja | mRNA | silny promotor owadów z genu Drosophila Actin 5c | konstytutywny | powszechnie stosowany w systemach ekspresji dla Drosophila. |
| Polihedryna | ogólna ekspresja | mRNA | silny promotor owadów z bakulowirusa | konstytutywny | powszechnie stosowany w systemach ekspresji dla komórek owadów. |
| CaMKIIa | ekspresja genu dla optogenetyki | mRNA | promotor zależnej od Ca2+/kalmoduliny kinazy białkowej II | swoisty | stosowany do ekspresji neuronów/OUN. Modulowane przez wapń i kalmodulinę. |
| GAL1, 10 | ogólna ekspresja | mRNA | drożdże sąsiadujące, rozbieżnie transkrybowane promotory | Indukowalne z galaktozą; tłumiące z glukozą | mogą być stosowane niezależnie lub razem. Regulowane przez GAL4 i GAL 80. |
| tef1 | ekspresja Ogólna | mRNA | promotor czynnika wydłużenia transkrypcji drożdży | konstytutywny | analogiczny do promotora ef1a ssaków. |
| GDS | ekspresja Ogólna | mRNA | silny promotor ekspresji drożdży z dehydrogenazy 3-fosforanowej aldehydu glicerolowego | konstytutywny | bardzo silny, zwany także TDH3 lub GAPDH. |
| ADH1 | ogólna ekspresja | mRNA | promotor drożdży dla dehydrogenazy alkoholowej i | tłumiony przez Etanol | pełna wersja jest silna z wysoką ekspresją. Okrojone promotory są konstytutywne o niższej ekspresji. |
| CaMV35S | ekspresja Ogólna | mRNA | silny promotor roślinny wirusa mozaiki kalafiora | konstytutywny | aktywny w dikotach, mniej aktywny w monokotach, z pewną aktywnością w komórkach zwierzęcych. |
| Ubi | ogólna ekspresja | mRNA | promotor roślin z genu ubikwityny kukurydzy | konstytutywny | daje wysoką ekspresję u roślin. |
| H1 | mała ekspresja RNA | shRNA | z promotora RNA ludzkiej polimerazy III | może mieć nieco niższą ekspresję niż U6. Może mieć lepszą ekspresję w komórkach neuronalnych. | |
| U6 | mała ekspresja RNA | shRNA | z ludzkiego U6 stosuje się również mały promotor jądrowy | konstytutywny | Mysi U6, ale może być mniej wydajny. |
promotorzy Prokariotyczni
| promotor | opis | wyrażenie | Uwagi dodatkowe | |
| T7 | transkrypcja in vitro/ ekspresja ogólna | promotor z bakteriofagu T7 | konstytutywny, ale wymaga polimerazy RNA T7. | gdy jest używany do transkrypcji in vitro, promotor napędza sensowny lub antysensowny transkrypt w zależności od jego orientacji na gen. |
| T7lac | wysokie poziomy ekspresji genów | promotor z bakteriofagów T7 i operatorów lac | nieistotna podstawowa ekspresja, gdy nie jest indukowana. Wymaga polimerazy RNA T7, która jest również kontrolowana przez operatora lac. Może być wywołany przez IPTG. | powszechnie spotykane w wektorach pET. Bardzo ściśle regulowane przez operatorów lac. Dobry do modulowania ekspresji genów poprzez zróżnicowane stężenia induktorów. |
| SP6 | transkrypcja in vitro/ ekspresja ogólna | promotor z bakteriofagu Sp6 | konstytutywny, ale wymaga polimerazy RNA SP6. | polimeraza SP6 ma wysoką procesywność. Gdy jest używany do transkrypcji in vitro, promotor napędza sensowny lub antysensowny transkrypt w zależności od jego orientacji na gen. |
| araBAD | ogólna ekspresja | promotor arabinozy metabolicznej operon | indukowany przez arabinozę i stłumiony katabolit represji w obecności glukozy lub przez kompetycyjne Wiązanie anty-induktora fukozy | słabszy. Powszechnie spotykany w wektorach pBAD. Dobry do szybkiej regulacji i niskiej ekspresji podstawowej; jednak nie nadaje się do modulowania ekspresji genów poprzez zróżnicowane stężenia induktorów. |
| TRP | wysoki poziom ekspresji genów | promotor z E. coli tryptofan operon | zostaje wyłączony z wysokim poziomem komórkowego tryptofanu. | |
| lac | wyrażenie ogólne | promotor z lac operon | konstytutywny w nieobecności Lac OPERON (lacI lub lacIq). Może być indukowany przez IPTG lub laktozę. | nieszczelny promotor o nieco słabej ekspresji. mutacja lacIq zwiększa ekspresję represora 10x, tym samym zaostrzając regulację promotora lac. Dobry do modulowania ekspresji genów poprzez zróżnicowane stężenia induktorów. |
| Ptac | ogólna ekspresja | Hybrydowy promotor lac i trp | regulowany podobnie jak promotor lac | zawiera -35 region z trpB i -10 region z lac. Bardzo ścisła Regulacja. Dobry do modulowania ekspresji genów poprzez zróżnicowane stężenia induktorów. Ogólnie lepsza ekspresja niż sama lac. |
| pL | wysoki poziom ekspresji genów | promotor z bakteriofagu lambda | może być regulowany temperaturą | często w połączeniu z wrażliwym na temperaturę represorem cI857. |
| T3 | transkrypcja in vitro/ogólna ekspresja | promotor z bakteriofagu T3 | konstytutywny, ale wymaga polimerazy RNA T3 | gdy jest stosowany do transkrypcji in vitro, promotor napędza sensowny lub antysensowny transkrypt w zależności od jego orientacji na gen |
chociaż ta lista jest doskonałym miejscem do rozpoczęcia, powyższe tabele nie zagłębiają się w tkanki lub promotory specyficzne dla rozwoju dostępne dla naukowców. Plazmidy są często wykorzystywane do celów terapeutycznych i w takich przypadkach ważne jest zidentyfikowanie odpowiednich promotorów specyficznych dla tkanek, jak opisano tutaj przez naukowców z NIH.
Uwaga: do napisania tego artykułu przyczynił się A. Max Juchheim.