ANNONSER:
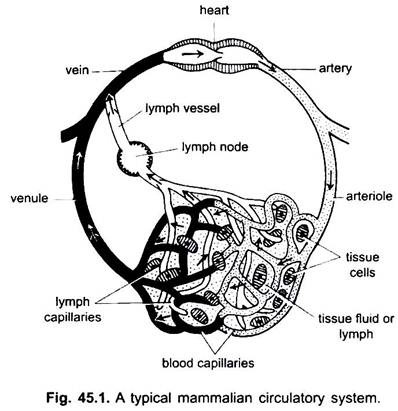
i sirkulasjonssystemet av vertebrater er det to systemer med utførlig forgreningsrør, som rammer gjennom hele kroppen og bærer væsker til vevet. De er et blodvaskulært system og et lymfesystem. Når begrepet sirkulasjonssystem brukes, refererer det bare til blodkarsystemet.
blodkarsystemet er et lukket system hos vertebrater; det har et kontraktilt hjerte og kontinuerlige rør kalt kar. Lymfesystemet er et åpent system med lymferom. Blodårene som fører blod bort fra hjertet er arterier, som deler seg i tynnere arterioler, forgrening i ekstremt tynne og små kapillærer.
veggen av en kapillær er laget av et enkelt lag av tasselated endotelceller. Hvert blodkar, inkludert hjertet, har en foring av endotelceller eller endotel. Blod kommer ikke i kontakt direkte med vevsceller.
ANNONSER:
Stoffer passerer fra og inn i kapillærene gjennom vævsfluidet som finnes i vevsrom mellom celler. Utveksling av stoffer mellom kroppens blod og vevsceller foregår gjennom kapillærveggene. Dette skyldes pinocytose i endotelceller i kapillærene.
under et elektronmikroskop viser endotelceller mange små vesikler som er invaginasjoner av plasmamembranene. Disse vesiklene beveger seg fra den ene siden av cellen og frigjør innholdet på motsatt side.
således gir blodet oksygen, næringsstoffer og hormoner til cellene for metabolisme, og cellene gir ut vann, karbondioksid og nitrogenholdig avfall i blodet for å bære dem til ekskretjonsorganene for rask eliminering.
kapillærene danner et nettverk i alle kroppsvev unntatt brusk og epitel. Fra kapillærene går blodet inn i tynne venules som kombinerer for å danne vener som bærer blod mot hjertet. Men noen årer (portal årer, nyre årer og lever vene) har kapillærer som er akkurat som de av arterier. Men alt blod passerer ikke gjennom kapillærene i venules.
ANNONSER:
Det er også noen gjennom kanaler mellom arterioler og venules som ligger i noen organer, for eksempel huden. Det er også arteriovenøse anastomoser mellom arterioler og venules funnet i sifrene. Funksjonen til slike forbindelser er ikke klart kjent, selv om det hevdes at de regulerer blodtrykk og sirkulasjon.
noen steder skjer utveksling av materiale mellom vev og blod gjennom tynnveggede rom eller sinusoider. I visse organer kan et blodkar danne et viklet nettverk av små blodkar kalt rete mirabile (nyrer, luftblære).
Deler Av Sirkulasjonssystemet:
ANNONSER:
i akkordater er sirkulasjonssystemet av lukket type. Denne typen finnes også i annelider (hvirvelløse dyr). En annen type er av åpen type der kapillærer ikke er funnet. Det finnes i bløtdyr og leddyr. Her strømmer blodet gjennom arterier i ulike organer, som passerer gjennom blodrom eller bihuler og deretter igjen i kar (årer) til hjertet.
sirkulasjonssystemet omfatter hjerte, arterier, vener, kapillærer og blod. Hjerte er et modifisert blodkar med muskulære vegger, som periodisk kontrakterer for å pumpe blodet inn i de ulike delene av kroppen gjennom bestemte kar.
Arterier og deres grener danner et arterielt system som bærer blodet fra hjertet. Årer og deres sideelver utgjør et venesystem som tar blodet fra kapillærene i arterier eller arterioler og bære det til hjertet.
Portalsystem:
i portalsystemet returneres blodet ikke direkte til hjertet, men det er et interposerende organ (lever eller nyre) i løpet av det returnerende blodet. Venen som bringer blod starter i kapillærene og slutter i kapillærene, den berørte venen virker både som afferent og efferent fartøy, de afferente karene slutter i kapillærene, akkurat som arterier, og blodet samles inn i systemiske årer.
alle virveldyr har et leverportalsystem hvor blod passerer inn i to sett med kapillærer i leveren. Lavere vertebrater og embryoer av høyere vertebrater har et nyreportalsystem også hvor blod passerer gjennom to sett med kapillærer i nyrene før de når hjertet. Kapillærene i hypofysen danner et hypofyseportalsystem som er et lite, men viktig system.
Lymfesystemet:
det finnes i akkordater unntatt syklostomer og bruskfisk. Det inkluderer lymfe og lymfekar. Lymfe er en vev væske funnet blant kroppens celler. Det er blodplasma minus røde blodlegemer og noen proteiner.
Lymfekapillærer som danner et nettverk av tynne blindendende kar, som samler lymf. Lymfekar er tynnveggede kar dannet ved forening av blodkarillærer. Disse tømmes i årer. Lymfeknuter finnes på lymfekar i pattedyr. Disse danner lymfocytter av blod som brukes til kroppsforsvar mot sykdommer.
Utvikling Av Hjerte Hos Vertebrater:
ANNONSER:
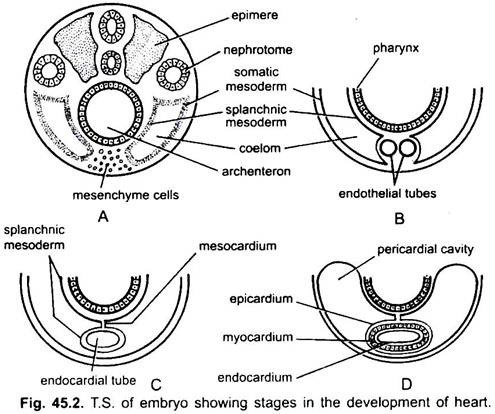
hjertet er et uparget organ, men dets opprinnelse er bilateralt. I et embryo danner mesenkymet en gruppe endokardiale celler under svelget. Disse cellene blir arrangert for å danne et par tynne endotelrør. De to endotelrørene smelter snart sammen for å danne et enkelt endokardialt rør som ligger i lengderetningen under svelget.
splanchnic mesoderm ligger under endoderm blir foldet i lengderetningen rundt endokardial røret. Dette tolags røret vil danne hjertet der splanchnic mesoderm tykkere for å danne et myokardium eller muskelvegg i hjertet og et ytre tynt epikardium eller visceral perikardium. Endocardial røret blir slimhinnen i hjertet kjent som endocardium.
Folder av splanchnic mesoderm møtes ovenfor for å danne en dorsal mesocardium som suspenderer hjertet i coelom. Snart en tverrgående septum er dannet bak hjertet som deler coelom i to kamre, en fremre perikard hulrom omslutter hjertet og en bakre bukhulen.
hjertet er et rett rør, men det øker i lengde og blir S-formet fordi endene er faste. Utseende av ventiler, innsnevring, partisjoner i hjertet og differensielle fortykkelser av veggene danner tre eller fire kamre i hjertet.
1. Single-Chambered Heart:
I Amphioxus (primitive akkordat), et sant hjerte er ikke funnet. En del av ventral aorta under svelget er muskuløs og kontraktil og fungerer som hjerte.
2. To-Kammer Hjerte:
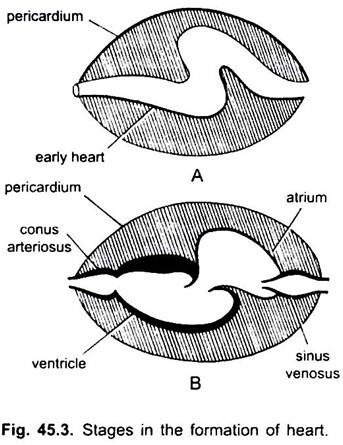
i syklostomer er det fire kamre arrangert i lineær rekkefølge – en tynnvegget sinus venosus, et litt muskulært atrium( auricle), en muskulær ventrikel og en muskuløs conus arteriosus eller bulbus cordis. Den ligger i kroppshulen der andre viscerale organer også er til stede.
av fire kamre svarer bare atrium og ventrikel til de fire kamrene (parrede atria og parrede ventrikler) av de høyere vertebrater. I utviklingen av hjertet har mange endringer skjedd.
Elasmobranchs:
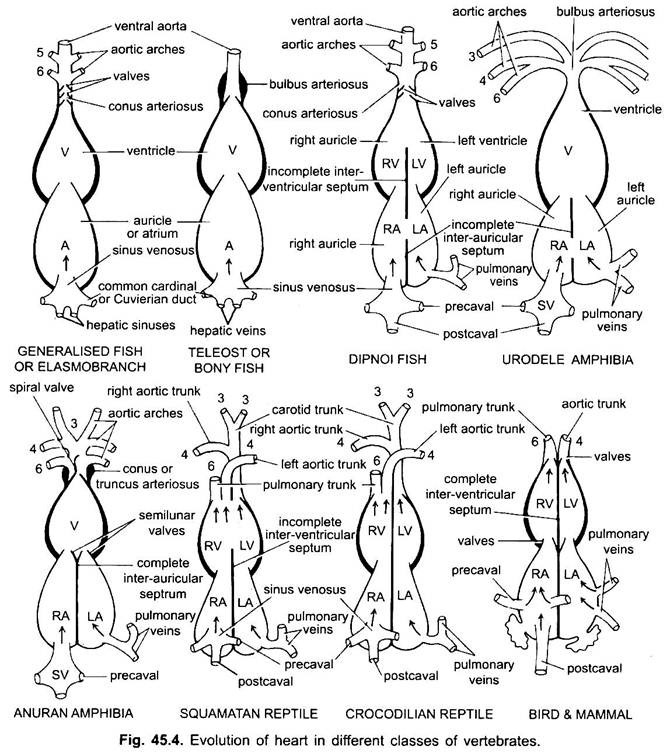
Unntatt Dipnoi, sirkulasjonssystemet i fisk fra cyclostomer til teleosts, går bare uoksygenert blod til hjertet, derfra pumpes det til gjellene, luftes og distribueres deretter til kroppen. Hjertet av bruskfisk er muskulært og dorsoventralt bøyd S-formet rør med fire rom i en lineær serie.
de er sinus venosus og atrium for å motta venøst blod, og en ventrikel og conus arteriosus for å pumpe dette blodet. Hjertet er et forgrenet venøst hjerte. Sinus venosus og conus arteriosus er tilbehørskamre. Atrium og ventrikel er sanne kamre, dermed er det et 2-kammerhjerte.
sinus venosus åpner anteriort inn i atrium gjennom sinu-atrial blenderåpning bevoktet av et par ventiler. Atrium ligger dorsal til ventrikel og åpner ventralt inn i ventrikel gjennom en atrio-ventrikulær blenderåpning bevoktet av et par ventiler. Den tykkveggede muskulære ventrikkelen åpnes i en smal conus arteriosus som inneholder ventiler i to serier.
hjertet er innelukket i perikard hulrom atskilt fra kroppshulen av en tverrgående septum. Conus pierces perikardiet og blir kontinuerlig med ventral aorta. Perikardial hulrom kommuniserer med kroppshulen gjennom to perforeringer i tverrgående septum.
Teleosts:
deres hjerte ligner på clasmobranchs. I teleosts er conus redusert og har et enkelt par ventiler. Den proksimale delen av ventral aorta nær conus blir sterkt forstørret og tykkvegget, kalt bulbus arteriosus. Det er elastisk og utvides på tidspunktet for ventrikulær sammentrekning. Hjertet er således 2-kammer med en enkelt blodsirkulasjon.
3. Trekammerat Hjerte:
Dipnoi:
i difoans deler en septum atriumet i et høyre og venstre kammer. Dette er korrelert med bruken av svømmeblæren som et åndedrettsorgan og representerer det første skrittet mot utviklingen av det dobbelte sirkulasjonssystemet hvor både oksygenert og uoksygenert blod kommer inn i hjertet og holdes separat.
Blod fra høyre auricle av lungfisken passerer inn i høyre ventrikel og pumpes deretter inn i den primitive lungelignende gassblæren av lungearterier som avgrener seg fra det sjette par aortabuer. Det oksygenerte blodet vender tilbake til venstre atrium ved hjelp av lungevenene som amfibier.
Amfibier:
i amfibier skifter dorsal atrium fremre til ventrikel. Sinus venosus åpner inn i høyre atrium dorsaltog ikke bakover. Atriet er helt delt inn i høyre og venstre kamre og har ingen foramen ovale i inter-auricular septum, som forblir åpen i dipnoans.
Dype lommer utvikles i ventrikulærhulen. Conus arteriosus deler seg i systemiske og lungekar med en spiralventil. I lunge mindre salamandere er inter-atrial septum ufullstendig og lungevenene er fraværende.
Reptilia:
i reptiler er hjertet ytterligere avansert. Atriumet er alltid helt separert i et høyre og venstre kammer, og i mange former er sinus venosus innlemmet i veggen til høyre atrium. Ventrikkelen er også delvis delt av en septum i de fleste reptiler, og i alligatorer og krokodiller er det helt tokammer.
dette betyr at oksygenert blod som kommer fra lungene til venstre side av hjertet, er i hovedsak skilt fra det ikke-oksygenerte blodet fra kroppen til høyre side. Således, i krokodilere, er de to blodtypene helt skilt, og nesten fullstendig i andre reptiler, men noe blanding forekommer i andre deler av sirkulasjonssystemet.
den embryonale conus arteriosus splittes i tre i stedet for to kar:
(i) Lungebue som frakter blod til lungene fra høyre side av ventrikkelen.
(ii) høyre systemisk aorta bærer blod fra venstre side av ventrikkelen til kroppen ved hjelp av høyre fjerde aortabuen.
(iii) venstre systemisk kommer fra høyre ventrikel til venstre fjerde aortabue.
ved kontaktpunktet med den systemiske aorta fra venstre ventrikel, selv hos krokodilere, er det en åpning mellom De To, kalt Foramen Av Panizzae, hvor det kan være noen blanding av de to blodtypene. Således representerer reptilisk hjerte overgangshjertet mot amfibisk hjerte – 2 komplette aurikler og 2 ufullstendige ventrikler med litt blanding av blod i høyre og venstre systemisk.
4. Firekammerat Hjerte:
Aves Og Mammalia:
hos fugler er ventrikkelen helt delt inn i to, slik at hjertet er fire kammer(2 aurikler og 2 ventrikler). Det er fullstendig separasjon av venøst og arterielt blod. Den systemiske aorta forlater venstre ventrikel og bærer blod til hodet og kroppen. Mens lungearterien forlater høyre ventrikel og bærer blod til lungene for oksygenering.
dermed er det dobbelt sirkulasjon der det ikke er blanding av blod på noe sted. Sinus venosus er helt innlemmet i høyre auricle, som mottar to precavals og en postcaval. Venstre auricle mottar oksygenert blod gjennom lungene, conus arteriosus er fraværende, pulmonal aorta oppstår fra høyre ventrikel, og enkelt systemisk aorta oppstår fra venstre ventrikel, og begge har ventiler på deres baser.
Modifikasjoner Av Aorta Buer I Virveldyr:
Embryonale Arterier:
når hjertet blir dannet i en virveldyr embryo, en blodåre kalt ventral aorta vises midt ventralt under svelget, som snart blir koblet til conus arteriosus. Den ventrale aorta som oppstår fra hjertet går fremover under svelget og deler fremre inn i et par ytre karotisarterier i hodet.
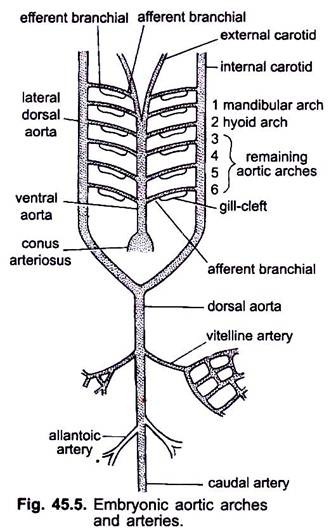
den ventrale aorta avgir 6 par laterale aortabuer ved ekvidistanse som går gjennom de viscerale buene. Hver aortabue har en ventral afferent grenarterie som bærer venøst blod til gjellen og en dorsal efferent grenarterie som tar oksygenert blod fra gjellen. De efferente forgreningsarteriene på hver side går sammen med den laterale dorsale aorta eller radix aorta som går inn i hodet som intern halspulsåren.
den første aortabuen er en mandibulær aortabue, den andre er en hyoid aortabue, de resterende kalles tredje, fjerde, femte og sjette aortabue. Den laterale dossal aorta smelter bak svelget for å danne en dorsal aorta som fortsetter midt dorsalt inn i halen som en kaudal arterie.
fra dorsal aorta oppstår parrede og uparede arterier som leverer ulike organer i kroppen. I et embryo med en eggeplomme, oppstår et par vitellinarterier fra dorsal aorta og forsyner eggeplommen. I embryoer av amnioter oppstår et par navlestreng eller allantoiske arterier fra dorsal aorta som leverer blod til allantois.
hos en voksen smelter vitellinarteriene sammen for å danne den viktigste mesenteriske arterien, hoveddelen av allantoiske arterier går tapt, men deres rester danner hypogastriske eller indre iliac arterier.
Aortabuer I Virveldyr:
i ulike voksne virveldyr ser det arterielle systemet ut til å være annerledes, men de er bygget på samme grunnleggende grunnplan. Forskjellen skyldes økt kompleksitet i hjertet på grunn av endring i respirasjon fra gjær til lunger. Det er en progressiv reduksjon i antall aortabuer i vertebrate-serien.
Cyclostomata:
I Petromyzon er det 7 par aortabuer. I andre syklostomer varierer disse fra 6 par I Myxin og 15 par I Eptatretus.
Fiskene:
selv om seks anses å ha vært det grunnleggende antall aortabuer for fisk. Dette tallet er redusert til fem, selv i haier og stråler med tap av det første paret, den mandibulære aortabuen eller den er representert av en efferent pseudobranchial arterie. I de fleste benfisk forsvinner både mandibulær (i) og hyoid (ii) aortabuene eller er mye redusert.
Hos Polypterus Og Dipnoi (lungefisk) er gjellene ikke godt utviklet. Så oppstår lungearterien fra den efferente delen av 6. bue på hver side som leverer blod til luftblæren eller lungen. I elasmobranchs og Dipnoi har hver bue en afferent og to efferente forgreningsarterier (dannet ved splitting) i hver gill. I benfisk har hver gill en afferent og en efferent arterie.
i tetrapoda aorta buer ikke bryte opp i afferente og efferente deler fordi sanne indre gjellene er fraværende. I alle tetrapoda forsvinner første og andre buer.
Amphibia:
her viser aortabuene endring på grunn av tap av gjær og utseende av lungene. I urodeles er det eksterne gjeller tilstede som åndedrettsorganer i tillegg til lungene. III, IV, v og vi aortabuer er til stede, men det femte paret er mye redusert I Sirene, Amphiuma og Necturus. Aorta buene er ikke brutt i de ytre gjellene i afferente og efferente deler, fordi grener som oppstår FRA IV, V og VI aorta buer danner kapillærer i de ytre gjellene.
den laterale dorsale aortaen mellom III og IV aortabuen fortsetter som en vaskulær forbindelse, ductus caroticus. VI aortabuen danner pulmo-kutan bue eller arterie på hver side tar blod til lungene og huden. Den beholder også en forbindelse med den laterale dorsale aorta kjent som en ductus arteriosus (kanal Av Botalli).
i larven av anuran (frosk tadpole) er arrangement av aorta buer som en voksen urodele på grunn av tilstedeværelse av gjær. Ved metamorfose, med tap av gjær, forsvinner I, II og V aorta buer helt, bare IIIrd, IVth og Vlth aorta buer er tilstede. Den laterale dorsale aorta mellom den tredje og fjerde aortabuen (ductus caroticus) forsvinner også. Dermed blir den tredje aortabuen sammen med en del av den ventrale aorta karotidbuen som bærer oksygenert blod til hodeområdet.
den fjerde aortabuen langs den laterale dorsale aorta danner den systemiske buen. Den sjette aortabuen blir pulmocutaneous buen leverer venøst blod til lungene og huden. Ductus arteriosus forsvinner under metamorfose. Således har voksne anuraner BARE III, IV OG vi aorta buer. Disse beholdes også av amnioter.
Reptilia:
i reptiler er gjellene fullt erstattet av lunger. BARE III, IV og vi aortabuer er til stede. Med den delvise separasjonen av ventrikkelen i to deler, er den distale delen av conus arteriosus og hele ventral aorta delt inn i tre kar, dvs. to aorta eller systemiske og en lunge. Høyre systemisk bue (IV) oppstår fra venstre ventrikel som bærer oksygenert blod til karotidbuen (III). Den venstre systemiske (IV) og pulmonale aortae (VI) tar sin opprinnelse fra høyre ventrikel. Den venstre systemiske bærer deoksygenert eller blandet blod til kroppen gjennom dorsal aorta.
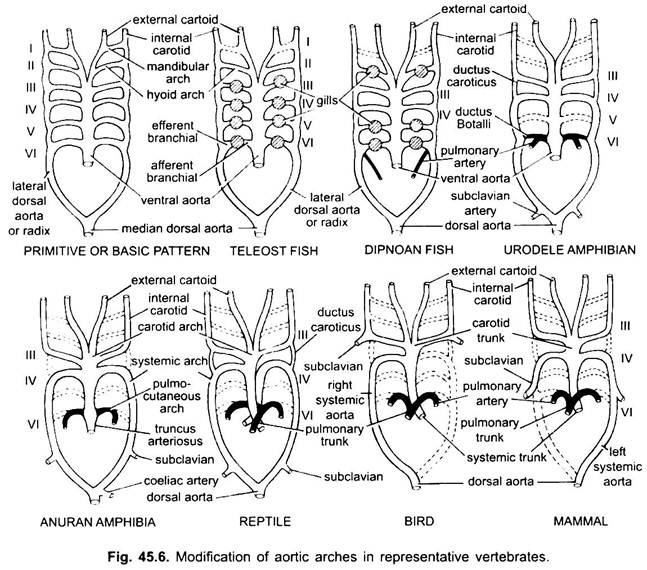
mens lungearterien tar deoksygenert blod til lungene. Ductus caroticus forsvinner, men det vedvarer i slanger og noen øgler (Uromastix). Ductus arteriosus forsvinner i de fleste reptiler selv om det vedvarer i redusert form I Sphenodon og noen skilpadder. På grunn av blanding av blod er reptiler kaldblodige dyr som fisk og amfibier.
Aves:
HOS fugler ER III, IV OG vi aortabuer til stede. De følger det generelle mønsteret av reptiler med noen forskjeller. Med den komplette delingen av ventrikkelen i to deler, har conus arteriosus og ventral aorta splittet for å danne to kar, systemisk aorta som oppstår fra venstre ventrikel og en pulmonal aorta fra høyre ventrikel.
Tredje aortabue med rester av laterale og ventrale aortaer danner karotidene som oppstår fra systemisk aorta. Fjerde aortabuen danner den systemiske aorta på høyre side bare. Den forener med den laterale aorta på sin egen side og danner dorsal aorta. En del av den fjerde aortabuen på venstre side danner den venstre subklaviske arterien, resten sammen med sin laterale dorsale aorta forsvinner. Den sjette aortabuen danner pulmonal aorta. Ductus caroticus og ductus arteriosus forsvinner.
Mammalia:
I pattedyr også III, IV og VI aoric buer vedvarer. Ventrikkelen er delt helt i to deler.
conus arteriosus og ventral aorta splittes for å danne to kar:
(i) en systemisk aorta som oppstår fra venstre ventrikel, og
(ii) en pulmonal aorta fra høyre ventrikel. Tredje aortabue med rester av lateral og ventral aortae danner karotidbuen.
Fjerde aortabuen danner den systemiske aorta på venstre side bare, mens på høyre side sin proksimale del danner en innominate og høyre subclavia arterie, resten sammen med sin laterale dorsal aorta forsvinner. Sjette aortabuen danner pulmonal aorta. Ductus arteriosus degenererer, men det vedvarer i noen til klekking eller fødsel i redusert form på venstre side som en tynn ligamentum arteriosum.
Venesystem:
Embryonale Vener:
i alle virveldyrembryoer er venesystemet enkelt og lignende. Årene er for det meste parret og symmetrisk arrangert. I embryoer uten en eggeplomme, dannes en sub – intestinal vene i splanchnic mesoderm under tarmen.
det løkker rundt anus og fortsetter bakover som:
(i) en kaudal vene inn i halen. I alle embryoer som har en plommesekk (enten de inneholder eggeplomme eller ikke), oppstår et par vitellinårer fra plommesekken og knytter seg til den bakre delen av det utviklende hjertet som blir sinus venosus. Faktisk er fusjonen av disse vitellinårene ansvarlig for hjertedannelsen i benfisk, reptiler og fugler. Hver vitellinvein i sin bakre ende forbinder den subintestinale venen dannet på samme måte som i embryoer som ikke har eggeplomme.
(ii) et par subcardinale vener oppstår mellom nyrene og knytter seg til kaudalvenen.
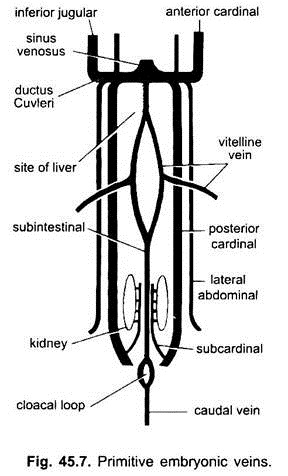
(iii) Parret fremre og bakre kardinalårer dannes, som bærer blodet fra henholdsvis hodet og bakre deler av kroppen. De fremre og bakre kardinalårene på hver side forenes for å danne en ductus Cuvieri eller felles kardinal vene som passerer innover gjennom tverrgående septum for å komme inn i sinus venosus.
i fisk og salamandere (urodeles) kommer en dårligere jugularvein fra den ventrale siden av hodet for å bli med i den vanlige kardinalvenen. Det har ingen homolog i andre vertebrater. I en amniotes kommer et par laterale eller ventrale abdominale vener fra kroppsveggen for å komme inn i de vanlige kardinalårene.
Fiskene:
Felles kardinal vene (kanal Av Cuvier) går inn i sinus venosus fra hver side og dannes ved sammensmelting av fremre og bakre kardinaler. Blod fra hodet samles av de fremre kardinaler, og blod fra nyrene og gonadene samles inn av bakre kardinaler.
også inn i kanalene Til Cuvier er de parede laterale bukårene som mottar blod fra kroppens vegg og parede vedlegg. Nyreportalsystemet består av den kaudale venen og to nyreportalår som ligger lateralt til nyrene som kapillariseres i nyrene.
Hepatisk portalsystem bærer blod fra mage og tarm og returnerer det til leveren, hvor det etter å ha passert gjennom serie sinusoider, går inn i sinus venosus ved hjelp av parede hapatiske årer.
i teleosts mangler de laterale bukårene, slik at blod fra subklaver, drenering av pectoral appendages, kommer inn i sinus venosus direkte, og blodet fra iliac vener, drenering av bekkenvedleggene, passerer inn i postkardinalene.
i dipnoans er en enkelt ventral abdominalvein tilstede, antagelig avledet fra en fusjon av laterale buk. Denne venen mottar blod fra ilacs ved hjelp av parede bekkenårer og går inn i Høyre kanal Av Cuvier. Videre, fra høyre postcardinal system, en ny vene, gjør postclaval sitt utseende som er av stor betydning hos høyere vertebrater. Det er forbundet med den kaudale venen og går fremover gjennom leveren til sinus.
i embryoer av amnioter er de laterale bukårene kjent som navlestreng eller allantoiske vener fordi de drenerer allantois. Vitellin og navlestreng er tapt ved fødselen, bare deres rester vedvarer.
Modifikasjoner Av Vener:
som i tilfelle av arterier, er venene til forskjellige vertebrater arrangert på samme grunnleggende plan. Eventuelle variasjoner vist av dem følger en sekvens i vertebrate-serien. I sin utvikling går venesystemet av høyere vertebrater gjennom stadiene sett i embryoer av lavere former.
1. Når leveren dannes den proksimale delen av vitellin-venene eller subintestinale vener danner leverenårene mellom leveren og hjertet, blir den distale delen av venstre vitellin-vene eller subintestinale vene den hepatiske portalvenen som danner sinusoider i leveren for å gi opphav til et hepatisk portalsystem som er tilstede i alle.
2. Anterior kardinal vener vedvarer som indre jugular vener.
3. Bortsett fra i fisk blir de vanlige kardinalårene de precavale venene som kommer inn i sinus venosus (amfibier, reptiler) eller høyre auricle (fugler, pattedyr). En subclavian vene dannes i hver forben som forbinder precaval venen.
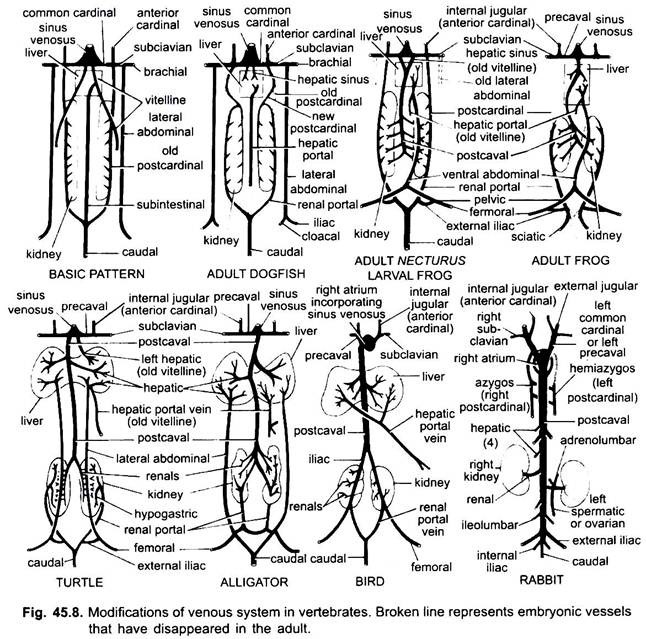
4. Den kaudale venen mister sine forbindelser med subintestinale og subkardinale vener, den fremre delen splittes i to grener som går sammen med de bakre kardinalårene i alle unntatt pattedyr.
5. Posterior kardinal årer vedvarer som sådan i fisk, men i andre hver bryter opp i to deler, en fremre del som forsvinner i amfibier, reptiler, og fugler, men i pattedyr høyre fremre del danner en azygos vene, den venstre kan danne en hemiazygos vene som mister sin forbindelse med precaval og tiltrer azygos vene ved en transversal anastomose; de bakre deler av bakre kardinal årer som har sluttet seg til halevenen, bli renal portal årer.
nyreportalvenen på hver side danner kapillærer i nyrene for å danne et nyreportalsystem som er komplett hos fisk, amfibier og reptiler, men blir mye redusert hos fugler, og er fraværende hos pattedyr fordi de bakre delene av bakre kardinalårene forsvinner hos pattedyr.
6. Nyreportalårene av fisk samler bare blod fra halen, men i amfibier, reptiler og fugler raner de også venene fra beina.
7. I noen fisk og alle tetrapoda de vitelline årer bli subcardinal vene for å danne en postcaval vene. I amfibier og reptiler strekker postkavalvenen seg til de bakre endene av nyrene, hos fugler går det sammen med nyreportalårene, og reduserer dermed nyreportalsystemet, hos pattedyr går postkavalvenen sammen med venene fra bena og halen, slik at nyreportalsystemet elimineres helt.
8. De to laterale abdominale venene vedvarer som sådan i fisk, men Fra Dipnoi oppover smelter de for å danne en fremre abdominalvene som knytter seg til leverportalvenen nær leveren, den fremre abdominalvenen forbinder dermed nyreportalen og leverportalsystemene. En iliac vene er dannet i hver hindlimb som forbinder den fremre abdominalvenen. Den fremre abdominalvenen begynner å miste sin betydning i reptiler, hos fugler er den modifisert for å danne epigastriske og coccygeo-mesenteriske vener, i pattedyr forsvinner den, unntatt I Takyglossus.
9. I luftpustende former vokser lungevenene ut fra venstre auricle og går inn i lungene.
10. Umbilical (allantoic) vener av embryoet forsvinner når lungene blir funksjonelle som åndedrettsorganer.
Tetrapoda:
venesystemet av amfibier ligner veldig mye på lungfisk, bortsett fra at abdominalvenen kommer inn i leverportalsystemet i stedet for sinus venosus. Fremre kardinaler vedvarer som indre jugular vener i alle voksne tetrapoder. Inferior jugular vener er fraværende. Vanlige kardinaler blir den fremre vena cavae eller precavals, som blir med sinus venosus i amfibier og reptiler.
hos fugler og pattedyr går precavaler direkte inn i høyre hjerte, fordi sinus venosus er fraværende. I mann, katt, etc., venstre precaval går tapt, slik at blodet på venstre side går inn i høyre precaval gjennom en brachio-cephalic gren. I lungeløse salamandere er lungevenene fraværende siden inter-atrial septum er ufullstendig.
postkardinalårene knytter seg til kaudalven bakover og precavaler fremover for å danne vanlige kardinaler i larvefrosk og Nekturus.
i reptiler, er det større utvikling av lungevenene og postcaval vene og reduksjon i betydningen av nyre portal system som bringer blod til nyrene fra den bakre delen av kroppen.
hos fugler er det to funksjonelle precavale vener dannet av foreningen av jugular og subclavian på hver side, og en komplett postkaval. Det mottar blod fra lemmer ved hjelp av nyreportaler, som passerer gjennom nyrer, men bryter ikke opp i kapillærene og er ikke sammenlignbare med nyreportaler av nedre vertebrater.
hos pattedyr kan det være en eller to precavale årer. Det er en enkelt postkavalvein, hvis embryologiske utvikling er svært komplisert. Alle kavale vener går direkte inn i høyre auricle, da sinus venosus absorberes i veggen av dette kammeret i hjertet i embryonalt liv. Det er ikke noe nyreportalsystem, selv om leverportalsystemet ligner mye på andre vertebrater.