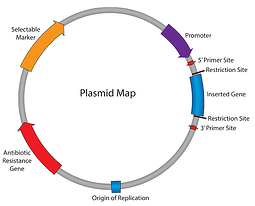
 Så langt i Vår Plasmids 101-serie har vi jobbet oss gjennom plasmid-kartet: antibiotikaresistens, replikasjonens opprinnelse og så videre. Opp til dette punktet kan vi replikere vårt plasmid og sørge for at cellene opprettholder det; det neste trinnet er å få plasmidet til å uttrykke vårt gen av interesse. Skriv inn promotoren-elementet som er ansvarlig for å starte transkripsjonen av innsatsen din I RNA.
Så langt i Vår Plasmids 101-serie har vi jobbet oss gjennom plasmid-kartet: antibiotikaresistens, replikasjonens opprinnelse og så videre. Opp til dette punktet kan vi replikere vårt plasmid og sørge for at cellene opprettholder det; det neste trinnet er å få plasmidet til å uttrykke vårt gen av interesse. Skriv inn promotoren-elementet som er ansvarlig for å starte transkripsjonen av innsatsen din I RNA.
i praksis beskriver begrepet «promoter» kombinasjonen av promotoren (rna-polymerasebindingssted) og operatører (responselementer). Promotorer er omtrent 100 til 1000 basepar lange og funnet oppstrøms av deres målgener. Sekvensen av promotorregionen styrer bindingen AV rna-polymerasen og transkripsjonsfaktorene, derfor spiller promotorer en stor rolle for å bestemme hvor og når ditt gen av interesse vil bli uttrykt.
rna-polymerasen (e)
RNA transkriberes FRA DNA ved hjelp AV EN rna-polymerase (RNAP). I bakterier gjøres dette av et enkelt enzym; eukaryoter har imidlertid flere polymeraser som hver er ansvarlige for en bestemt delmengde Av Rna. For å oppnå denne spesifisiteten kan eukaryote RNAP gjenkjenne og binde seg til bestemte promotorelementer. Dette betyr at promotoren som er tilstede i plasmid-ryggraden må være kompatibel med typen RNA som må gjøres: hvis du vil ha mRNA (for genuttrykk), må du bruke EN rnap II-promotør, mens små Rna (for eksempel shRNA) transkriberes fra RNAP III-promotørene. Dette innlegget har arrangører for generell rnap II og rnap III transkripsjon; derimot, ved hjelp av virale LTRs som RNAP II arrangører er ofte ansatt i lentiviral og retrovirale konstruksjoner, og vi vil diskutere disse i en fremtidig innlegg på viral vektor deler.
Promotorspesifisitet
Bortsett fra å velge en promotør basert på TYPE rna-transkripsjon, må du også sørge for at plasmidet ditt har en promotør som er egnet til å jobbe i vertsorganismen din. Fordi transkripsjonsmaskiner varierer mellom celletyper eller organismer, må promotorer være like variable. Bakterielle promotorer virker bare i prokaryote celler og vanligvis bare i samme eller nært beslektede arter som de ble avledet fra. På samme måte krever de forskjellige eukaryote celletypene (pattedyr, gjær, planter, etc) unike promotorer, og det er svært lite crossover. Generelt sett er promotorer i bakterier mindre varierte og komplekse, med færre deler enn de i eukaryote celler. Noen promotorer er konstitusjonelt aktive og hele tiden, mens andre er mer nøye kontrollert. Regulerte promotorer kan bare handle i visse vev eller på bestemte tidspunkter i utvikling, eller det kan være måter å slå dem på eller av på vilje med kjemikalie, varme eller lys. I cellen styres promotorer selv av fortsatt andre regulatoriske faktorer: forsterkere, grenseelementer, isolatorer og lyddempere; det kan imidlertid forekomme noe «lekkasje» av transkripsjon. Dette er normalt ikke et stort problem for celler, men det kan forvirre forskningsresultater eller til og med drepe cellene dine hvis ditt gen av interesse er giftig. For å bekjempe dette har forskere skapt syntetiske promotorer, som vanligvis inneholder en kombinasjon av andre promotorelementer, og har en tendens til å være tettere regulert.
Vanlige promotører for eukaryoter og prokaryoter
vi har tatt med to referansetabeller nedenfor som viser noen av de vanligste bakterie-og pattedyrpromotørene. Disse listene er på ingen måte uttømmende, men bør være et godt sted å starte når du prøver å plukke din perfekte arrangøren!
Eukaryotiske Promotører
| Promotør | Primært brukt til | rna-transkripsjon | Beskrivelse | Uttrykk | Flere hensyn |
| CMV | Generelt uttrykk | mRNA | sterke pattedyr uttrykk promotor fra humant cytomegalovirus | Konstitutiv | kan inneholde en forsterker region. Kan være stille i enkelte celletyper. |
| EF1a | Generelt uttrykk | mRNA | Sterkt pattedyruttrykk fra menneskelig forlengelsesfaktor 1 alfa | Konstitutiv | har en tendens til å gi konsekvent uttrykk uavhengig av celletype eller fysiologi. |
| SV40 | Generelt uttrykk | mRNA | Pattedyruttrykkspromotor fra simian vacuolating virus 40 | Konstitutiv | kan inneholde en forsterker. |
| PGK1 (menneske eller mus) | Generelt uttrykk | mRNA | Pattedyrpromotor fra fosfoglyseratkinasegen. | Konstitutiv | Utbredt uttrykk, men kan variere etter celletype. Tendens til å motstå promoter ned regulering på grunn av metylering eller deacetylering. |
| Ubc | Generelt uttrykk | mRNA | Pattedyrpromotor fra det humane ubiquitin C-genet | Konstitutiv | som navnet tilsier, er denne promotoren allestedsnærværende. |
| human beta actin | Generelt uttrykk | mRNA | Pattedyrpromotor fra beta actin-genet | Konstitutiv | Allestedsnærværende. Kylling versjon er ofte brukt i promoter hybrider. |
| CAG | Generelt uttrykk | mRNA | Sterk hybrid pattedyr arrangøren | Konstitutiv | Inneholder CMV enhancer, kylling beta aktin arrangøren, og kanin beta-globin skjøte akseptor. |
| TRE | Generelt uttrykk | mRNA | tetracyklinresponselementpromotor | Induserbar Med Tetracylin eller dets derivater. | Inneholder Vanligvis en minimal promoter med lav basal aktivitet og flere tetracyklinoperatører. Transkripsjon kan slås på eller av avhengig av hva tet transactivator brukes. |
| UAS | Generelt uttrykk | mRNA | Drosophila arrangøren conaining Gal4 bindende steder | Spesifikke | Krever Tilstedeværelse Av Gal4 genet for å aktivere arrangøren. |
| Ac5 | Generelt uttrykk | mRNA | Sterk insekt promoter Fra Drosophila Actin 5c genet | Konstitutiv | Vanligvis brukt i uttrykk systemer For Drosophila. |
| Polyhedrin | Generelt uttrykk | mRNA | Sterk insektpromotor fra baculovirus | Konstitutiv | Vanligvis brukt i uttrykkssystemer for insektceller. |
| CaMKIIa | Genuttrykk for optogenetikk | mRNA | Ca2+/calmodulinavhengig proteinkinase II-promotor | Spesifikk | Brukes til neuronal / CNS-uttrykk. Modulert av kalsium og calmodulin. |
| GAL1, 10 | Generelt uttrykk | mRNA | Gjær tilstøtende, divergent transkribert promotorer | Induserbar med galaktose; undertrykkende med glukose | kan brukes uavhengig eller sammen. Regulert AV GAL4 OG GAL 80. |
| TEF1 | Generelt uttrykk | mRNA | gjærtranskripsjon forlengelse faktor promoter | Konstitutiv | Analog til pattedyr EF1a promoter. |
| GDS | Generelt uttrykk | mRNA | sterk gjær uttrykk promoter fra glyceraldehyd 3-fosfordehydrogenase | Konstitutiv | Veldig sterk, Også kalt TDH3 ELLER GAPDH. |
| ADH1 | Generelt uttrykk | mRNA | Gjærpromotor for alkoholdehydrogenase I | undertrykt av etanol | full lengde versjon er sterk med høyt uttrykk. Avkortede promotorer er konstitutive med lavere uttrykk. |
| CaMV35S | Generelt uttrykk | mRNA | Sterk plantepromotor Fra Blomkålmosaikkviruset | Konstitutiv | Aktiv i dikoter, mindre aktiv i monokoter, med noe aktivitet i dyreceller. |
| Ubi | Generelt uttrykk | mRNA | Plantepromotor fra mais ubiquitingenet | Konstitutiv | Gir høyt uttrykk i planter. |
| H1 | små rna uttrykk | shRNA | fra human polymerase III rna promoter | Konstitutiv | Kan ha noe lavere uttrykk Enn U6. Kan ha bedre uttrykk i nevronceller. |
| U6 | lite RNA-uttrykk | shRNA | fra den menneskelige u6 liten kjernefysisk promoter | Konstitutiv | Murine U6 brukes også, men kan være mindre effektiv. |
Prokaryotiske promotorer
| Promotør | Primært brukt for | Beskrivelse | Uttrykk | |
| T7 | in vitro transkripsjon / generelt uttrykk | Promoter Fra t7 bakteriofag | Konstitutiv, men krever T7 RNA-polymerase. | når den brukes for in vitro transkripsjon, driver arrangøren enten sense eller antisense transkripsjon avhengig av sin orientering til genet. |
| T7lac | Høye nivåer av genuttrykk | Promoter Fra t7 bakteriofag pluss lac-operatører | Ubetydelig basal ekspresjon når den ikke er indusert. Krever T7 RNA-polymerase, som ogsa styres av lac-operatoren. Kan induseres av IPTG. | Vanligvis funnet i pET vektorer. Veldig tett regulert av lac-operatørene. Bra for modulerende genuttrykk gjennom varierte induktorkonsentrasjoner. |
| Sp6 | in vitro transkripsjon/ generelt uttrykk | Promoter Fra Sp6 bakteriofag | Konstitutiv, men krever SP6 RNA-polymerase. | sp6 polymerase har en høy prosessivitet. Når den brukes for in vitro transkripsjon, driver arrangøren enten sense eller antisense transkripsjon avhengig av sin orientering til genet. |
| araBAD | Generelt uttrykk | Promotor av arabinosemetabolismen operon | Induserbar ved arabinose og undertrykt katabolitt undertrykkelse i nærvær av glukose eller ved konkurransedyktig binding av anti-induktoren fucose | Svakere. Vanligvis funnet i pbadvektorer. Bra for rask regulering og lav basal ekspresjon; men ikke godt egnet for modulerende genuttrykk gjennom varierte induktorkonsentrasjoner. |
| trp | Høye nivåer av genuttrykk | Promoter Fra E. coli tryptofan operon | Undertrykkende | blir slått av med høye nivåer av cellulær tryptofan. |
| lac | Generelt uttrykk | Promoter fra lac operon | Konstitutiv i fravær av lac repressor (lacI eller lacIq). Kan induseres AV iptg eller laktose. | Lekk promoter med noe svakt uttrykk. lacIq mutasjon øker ekspresjon av repressor 10x, og dermed stramme regulering av lac promoter. Bra for modulerende genuttrykk gjennom varierte induktorkonsentrasjoner. |
| Ptac | Generelt uttrykk | Hybrid promoter av lac og trp | Regulert som lac promoter | Inneholder -35 region fra trpB og -10 region fra lac. Veldig stram regulering. Bra for modulerende genuttrykk gjennom varierte induktorkonsentrasjoner. Generelt bedre uttrykk enn lac alene. |
| pL | Høye nivåer av genuttrykk | Promoter fra bakteriofag lambda | kan være temperaturregulerbar | ofte parret med den temperaturfølsomme cI857-repressoren. |
| T3 | in vitro transkripsjon/generelt uttrykk | Promotør Fra T3 bakteriofag | Konstitutiv, men krever T3 RNA-polymerase | når den brukes til in vitro transkripsjon, driver promotoren enten sense-eller antisense-transkripsjonen avhengig av retningen til genet ditt |
selv om denne listen er et flott sted å starte, tabellene ovenfor ikke dykke inn i vev eller utvikling-spesifikke arrangører tilgjengelig for forskere. Plasmider blir ofte satt til terapeutisk bruk, og i slike tilfeller er det viktig å identifisere de riktige vevsspesifikke promotorene som beskrevet av forskere ved NIH her.
Merk: A. Max Juchheim bidro til skrivingen av denne artikkelen.