Betydning
det er en økende interesse for circrna på grunn av deres implikasjon i mange biologiske prosesser og sykdommer i tillegg til deres biomarkørpotensial. De er hovedsakelig oppdaget av tilstedeværelsen av leser kartlegging deres backsplicing veikryss. Ikke desto mindre er circrna ikke lenger de eneste transkripsjonene som inneholder et slikt veikryss siden nyere studier har avslørt at sirkulære Dnaer er vanlige og kan transkriberes, noe som resulterer i transkripsjoner som vil etterligne et circRNA-signal. Derfor kan denne nye typen kimære transkripsjon endre måten circRNA analyse blir gjort og påvirke noen av resultatene som allerede er rapportert.
Er Sirkulære Rna de Eneste Kimære Transkripsjonene?
Sirkulært RNA (circRNAs) ble gjenoppdaget for noen år siden som ikke-kanonisk spleiset RNA former til stede i forskjellige organismer, inkludert mennesker (Salzman et al., 2012; Jeck et al., 2013). De er kovalent lukkede transkripsjoner dannet gjennom EN rna-spleisingshendelse, hvor en spleisedonor av en nedstrøms ekson knytter seg til en oppstrøms spleiseacceptor som fører til kovalent lukkede transkripsjoner som er preget av tilstedeværelsen av et back-spleisingsknutepunkt som gjør circRNAs skiller seg fra deres lineære motstykker (Figur 1a) (Zhang et al.( 2016; Wilusz, 2018).
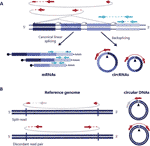
Figur 1 Deteksjon av både sirkulært RNA og DNAs er basert på leser som spenner over krysset. (A) Backspliced sammenføyning mellom eksoner og kanonisk ekson sammenføyning er avbildet med henholdsvis ikke-kontinuerlige og kontinuerlige linjer som gir opphav til circrna og mrna. Sammenkoblede endelesninger som spenner over de bakre kryssene, vises i rødt, og sammenkoblede endelesninger som samsvarer med et lineært transkripsjon, vises i blått. (B) i Likhet med circRNAs, blir sirkulære DNAs (eccDNAs) detektert basert på strukturelle lesevarianter i samsvar med en sirkulasjonshendelse avbildet i rødt .
siden gjenoppdagelsen har det vitenskapelige samfunn trukket oppmerksomheten mot circRNAs og har undersøkt deres engasjement i flere cellulære prosesser i helse og sykdom (Haque and Harries, 2017), deres potensielle rolle som biomarkører (Abu Og Jamal, 2016) og deres regulatoriske funksjoner (Floris et al., 2016). Circrna er nå kjent for å være rikelig og stabil i cytosol og kjernen (Salzman et al., 2012; Jeck et al., 2013; Li et al., 2015) og har også blitt funnet fri i biofluider (Bahn et al., 2015; Memczak et al., 2015; Chen et al., 2018) og i ekstracellulære vesikler (Kyoung Mi et al., 2017). Biomarkørpotensialet til circRNAs har blitt intensivt studert, faktisk har det blitt publisert mange case-control-studier som søker differensielt uttrykte circRNAs som kan være biomarkører av forskjellige sykdommer. Til dags dato har circrna vært involvert i flere sykdommer, inkludert kreft (Kristensen et al., 2017; Arnaiz et al., 2018), nevrologiske lidelser (Akhter, 2018), kardiovaskulære sykdommer (Aufiero et al., 2019) og immunrelaterte sykdommer (Iparraguirre et al., 2017; Liu et al., 2019). Samtidig, å få til å forstå deres biogenese, egenskaper, funksjoner og implikasjoner i menneskelig biologi forbli som åpne spørsmål for forskere på feltet.
selv om funksjonen til de fleste av circRNAs forblir ukjent, har det vist seg at noen circRNAs kan fungere som microRNA svamper, regulere microRNA nivåer og deres aktivitet (Hansen et al., 2013; Memczak et al., 2013; Zheng et al., 2016). De er involvert i genuttrykksregulering ved å regulere transkripsjonen av foreldrenes gener, og konkurrerer med de lineære spleising eller svampende proteiner (Ashwal-Fluss et al., 2014; Li et al., 2018). Interessant, ribosom profilering studier har nylig vist at circRNAs kan også oversettes både in vitro og in vivo (Legnini et al., 2017; Jan et al., 2017).
hovedtrekk ved circRNAs, og ansvarlig for de fleste av deres spesielle egenskaper, er deres sirkularitet. Derfor, i tillegg til å oppdage deres karakteristiske back-skjøtes veikryss, teste sirkularitet av disse molekylene, er en av de viktigste punktene i hver circRNA studie. Likevel har mange studier basert sin oppdagelse av circrna på totalt RNA og kan dermed ha tolket noen lineære kimære transkripsjoner som circrna, noe som resulterer i falske positive circRNA-deteksjoner. For å omgå dette problemet har de fleste studier bekreftet sirkulariteten til transkripsjonene funnet av total RNA-seq ved Hjelp Av Rnase R, Northern blot eller elektroforetiske metoder (Jeck og Sharpless, 2014). Imidlertid har disse sirkulasjonsvalideringene også noen ganger avslørt transkripsjoner som synes å være lineære, i stedet for sirkulære som bekrefter at deteksjon av sirkrna som starter fra totalt RNA, kan føre til noen falske positiver. Disse falske positivene har blitt tilskrevet tekniske artefakter eller transkripsjoner avledet fra uvanlige hendelser som eksonduplikasjoner eller transplantasjonshendelser (Jeck og Sharpless, 2014; Szabo og Salzman, 2016). Når det er sagt, har muligheten til å ha funnet sanne, biologisk aktive og funksjonelle lineære transkripsjoner som inneholder en sekvens som tilsvarer et backsplicing-kryss (fra nå av kalt kimære lineære transkripsjoner), blitt noe oversett fordi en kilde til slikt lineært RNA ikke har vært kjent for friske celler.
Sirkulære Dnaer Som Kilde Til Kimære Lineære Transkripsjoner
Det meste av det menneskelige genomet er organisert i lineære kromosomer, men noen unntak har lenge blitt akseptert som mitokondrielt DNA og kromosomavvik som DNA-sirkler som bærer onkogener (f. eks. doble minutter) (Benner et al ., 1991; Nathanson et al., 2014; Turner et al., 2017) og ringkromosomer (Tü et al., 2004). Det var ikke før nylig at forskjellige sirkulære DNAs som microDNAs (Shibata et al., 2012) eller extrachromosomal sirkulær DNAs (eccDNAs) ble funnet å også oppstå fra store deler av ulike eukaryote genomer inkludert human og gjær(Mø et al ., 2015; Mikkelsen et al., 2017; Mø et al., 2018).
Sirkulære Dnaer dannes når to ender av et lineært DNA er felles sammen, noe som resulterer i et kryss som ligner det bakre krysset på sirkrnaer som ofte kalles brytpunktskryss som oppdages basert på strukturelle lesevarianter i samsvar med en sirkulasjonshendelse (Gresham et al., 2010; Mø et al., 2018; Prada-Luengo et al. 2019) (Figur 1b). De varierer vanligvis fra hundre baser til megabase sirkler og kan inneholde fulle eksoner og gener (Shibata et al ., 2012; Mø et al., 2015; Mikkelsen et al., 2017; Geir et al., 2017; Mø et al., 2018) og mens noen regioner av genomet er mer vanlig funnet på sirkulært DNA (Sinclair og Guarente, 1997;; Mø et al., 2016; Geir et al., 2017; Mø et al., 2018), ser det ut til at de fleste sirkulære DNA forekommer tilfeldig (Shibata et al., 2012; Mø et al., 2015; Mikkelsen et al., 2017; Mø et al., 2018).
Interessant, i et nylig papir Mø et al. identifisert tusenvis av eccdna i leukocytter og muskelceller i friske kontroller. Med ideen om å undersøke om eccdna kunne transkriberes, ble et mRNA-bibliotek også sekvensert fra muskelvev og analysert for transkripsjonshendelser over brytpunktskrysset av det oppdagede eccDNA å finne flere kamper (Mø et al ., 2018). Dette funnet antyder at sirkulært DNA i sunt vev transkriberes, noe som gir opphav til lineære og polyadenylerte transkripsjoner som vil bære en sekvens som tilsvarer backsplicing-sekvensen av circRNAs (Mø et al., 2018) (Figur 2).
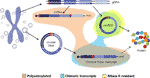
Figur 2 Grafisk fremstilling av forskjellige transkripsjoner som stammer fra enten genomisk DNA ELLER sirkulært DNA. Exons er farget i lilla og backspliced veikryss eller kimære veikryss er vist i rødt. Polyadenylerte, kimære kryss som inneholder Og rnase R-resistente transkripsjoner er uthevet i henholdsvis oransje, blå og gul.
det transkripsjonelle beviset på sirkulære Dnaer, sammen med deres overflod, fører oss til å foreslå at sirkulære Dnaer kan være en naturlig kilde til en betydelig mengde lineære Rna som bærer kimære veikryss. I mange tilfeller, disse kimære veikryss kan være umulig å skille fra backsplicing veikryss av circRNAs, og derfor kan de være forvirrende faktorer i circRNA studier. I de følgende avsnittene vil vi forklare dataene som støtter dette forslaget.
sirkrna-Gjenkjenning: Alt Som Glitter Er Ikke Gull
som tidligere introdusert, dannes circRNAs om en ikke-kanonisk spleising hendelse kalt backsplicing. Transkripsjoner som følge av denne backsplicing hendelsen har en kovalent lukket sløyfe struktur med verken 5′ -3 ‘ polaritet, eller en polyadenylated hale og enda viktigere, de er preget av tilstedeværelsen av en kryptert exon orden i forhold til den lineære transkripsjon (Zhang et al .( 2016; Wilusz, 2018). Denne krypterte exon order blir tydelig i backspliced junction som forbinder en 5 ‘nedstrøms sekvens med en oppstrøms 3’ sekvens. Dermed utnytter alle circRNA-deteksjonsalgoritmer tilstedeværelsen av de bakre skjøtede kryssene som en diagnostisk funksjon for identifisering av circRNA (Figur 1a).
Ulike metoder har blitt tilpasset for påvisning av disse back-skjøtes veikryss. Kommersielle arrays inneholder sonder rettet mot disse backspliced regioner har vært mye brukt i biomarkør screening studier (Iparraguirre et Al., 2017; Liu et al., 2017; Sui et al., 2017; Li et al., 2018). Den etterfølgende valideringen er ofte også basert på forsterkning av backsplicingkryssene ved hjelp av divergerende primere (Panda og Gorospe, 2018). Mange andre papirer har gjennomført en høy gjennomstrømning sekvensering analyse som overvinner en av de viktigste begrensningene i arrays tillater å oppdage ikke bare annotert circRNAs men også de novo rna sirkularisering hendelser fra genomiske regioner der ingen circRNA ble annotert av tidligere studier. Flere bioinformatiske rørledninger er utviklet for påvisning av circRNAs I RNA-Seq datasett, men alle er basert på tilstedeværelsen av leser som krysser over rygg-spleisekryssene, og å finne den mest pålitelige er fortsatt en utfordring for bioinformatikere (Hansen et al., 2016; Hansen, 2018; Prada-Luengo et al., 2019).
To hovedtilnærminger kan følges for påvisning av circrna i RNA-Seq-data. For det første er mange circRNA RNA-seq-studier basert På rnase r-behandlede prøver for å tømme alle lineære Rna før sekvensering. Selv om denne tilnærmingen er spesielt designet for circRNA-deteksjon, er Det verdt å merke Seg At rnase R-nedbrytning er variabel, at det er sjeldne tilfeller Av rnase r-resistente lineære rna og rnase r-sensitive circrna (Szabo and Salzman, 2016) og at denne behandlingen krever en høy RNA-inngang som kan begrense for noen vev. Andre circRNA-studier velger å sekvensere enten totalt, ribosomalt utarmet (ribo -) ELLER ikke-polyadenylert (polyA-) RNA, hvor både lineære Og sirkulære Rna kan bli funnet (Salzman et al., 2012; Memczak et al., 2013; Broadbent et al., 2015; Lu et al., 2015; Memczak et al., 2015). Denne tilnærmingen unngår bruken Av RNase R, noe som reduserer rna-mengden som trengs for sekvensering og tillater å studere uttrykket av andre Typer Rna fra samme datasett. Det har vist seg at med en god sekvenseringsdybde og kvalitet og en nøye dataanalyse kan sanne sirkrnaer detekteres fra total RNA-sekvensering (Wang et al., 2017), men i denne andre tilnærmingen er det nødvendig med en senere sirkulasjonsbekreftelse for circRNAs.
med oppdagelsen av lineære kimære Rna transkribert fra sirkulære Dnaer, er sirkrna ikke lenger de eneste transkripsjonene med kimære veikryss. Derfor er det av største betydning å merke seg at mens den første tilnærmingen vil betydelig berike RNA-prøven i circRNAs, slik at de fleste av de oppdagede kimære kryssene vil korrespondere med sanne circRNAs, kan den andre overvurdere antall circRNA-transkripsjoner ved å tildele circRNAs signalet som kommer fra både circRNAs og de lineære kimære transkripsjonene transkribert fra sirkulære DNAs. Følgelig, tar hensyn til sameksistens av circRNAs og lineære kimære transkripsjoner, behovet for sirkularitet tester og funksjonalitet analyser gevinster betydning og spesiell forsiktighet bør tas om ikke bare eksperimentelle, men også beregningsmetoder for å unngå feil kimære transkripsjoner fra sirkulære DNAs med circRNAs dannet av backsplicing.
Diskusjon
circRNA-feltet er fortsatt på et tidlig stadium, men circrna har allerede vist seg å være forbløffende molekyler, involvert i mange prosesser, med et stort biomarkørpotensial, og det kan også endre måten vi forstår transkripsjons-og oversettelsesprosessene på. Av disse grunnene får de oppmerksomhet, og circRNA-feltet er for øyeblikket et AV DE mest aktive rna-forskningsfeltene. Det er imidlertid fortsatt mange konflikter ,kontroverser og åpne spørsmål (Li, 2019) som må diskuteres.
i denne rapporten, og i lys av de siste fremskrittene i det sirkulære DNA-feltet, ønsker vi å påpeke transkripsjonen fra ekstrakromosomalt sirkulært DNA som en av de viktigste naturlige kildene til lineære transkripsjoner med baksplittede signaler som kan forstyrre circRNA-data (Mø et al., 2018). Fra nå av, bortsett fra de tekniske artefakter, duplikasjoner og transplantasjonshendelser som kan føre til falske positiver i circRNA-deteksjonen, bør vi også ta hensyn til eksistensen av denne nye typen kimære transkripsjoner. Derfor er sirkulasjonstester og funksjonelle analyser viktigere enn noensinne.
i alle fall bør disse kimære lineære transkripsjonene ikke bare betraktes som en forvirrende faktor for circRNA-studier. Til tross for de tekniske implikasjonene for circRNA-karakteriseringen, legger eksistensen av disse circRNA-lignende kimære lineære rna-molekylene fra eccdna til en ny type molekyl til den stadig voksende listen Over rna og utvider vår visjon om kompleksiteten til transkriptomet og dets regulering. Videre kan disse lineære rna-molekylene som kommer fra eccDNA også presentere funksjoner som ligner på circRNA, inkludert regulatoriske funksjoner eller potensialet som skal oversettes. Genprodukter fra eccDNA-transkripsjoner kan potensielt bidra til fenotypen av somatiske celler og vev som rapportert i gjær (Gresham et al ., 2010; Demeke et al., 2015). Men i dette nascent feltet er det behov for mer data og forskning for å begynne å skrape overflaten av isfjellet.
Forfatterbidrag
LI, IP-L, BR og DO skrev papiret.
Finansiering
denne studien er finansiert Av Instituto De Salud Carlos III gjennom prosjektet » PI17/00189 «(Medfinansiert Av European Regional Development Fund/European Social Fund)»Investing in your future»). IP-L OG BR ble støttet av dansk Råd For Uavhengig Forskning, 6108 – 00171B OG LI ble støttet av Utdanningsdepartementet I Den Baskiske Regjeringen .
Interessekonflikter
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.