ANNONSER:
i denne artikkelen vil vi diskutere om fordøyelsessystemet av vertebrater ved hjelp av egnede diagrammer.
Embryonale Fordøyelseskanalen:
Archenteron:
den embryonale archenteron blir slimhinnen i den voksne fordøyelseskanalen og av alle dets derivater. Splanchnic mesoderm legger til lag av bindevev og glatte muskler rundt archenteron. Ectodermal invaginering av hodet danner stomodaeum som fører inn i munnhulen, og en lignende mid-ventral ectodermal invaginering danner proctodaeum, som fører inn i hindgut.
ANNONSER:
stomodeum blir det voksne bukkalhulen og gir opphav til tannemalje, epitelbelegg av tungen, kjertler, f. eks. slimete, gift og spytt, etc. Og rathkes pose med fremre hypofyse. Proctodaeum danner enten en liten terminal del av cloaca i nedre vertebrater og endetarm hos pattedyr.
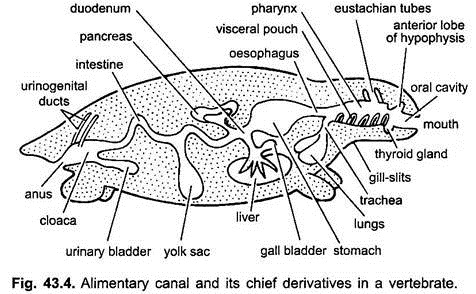
fordøyelseskanalen i embryoer fra mage til cloaca er festet til den dorsale kroppsveggen ved en dobbel fold av peritoneum, kalt dorsal mesenteri, og til ventral kroppsveggen av en ventral mesenteri. Hos voksne vedvarer dorsal mesenteri, men ventral mesenteri forsvinner, slik at bare i lever-og urinblæren.
Fordøyelseskanal Av Voksen:
fordøyelseskanalen skiller seg for forskjellige funksjoner i følgende regioner-munn, bukkal hulrom, svelg, øsofagus, mage, tynntarm, tykktarm og cloaca. Etter utvekster oppstår fra fordøyelseskanalen – orale kjertler, rathkes pose, skjoldbruskkjertel, gill-clefts, tympanisk hulrom, tymus og andre kjertler av gill-clefts, luftrør, lunger, svømmeblære, lever, bukspyttkjertel, eggeplomme og urinblære.
ANNONSER:
Histologi:
veggen av fordøyelseskanalen er laget av fire konsentriske lag.
De er:
(i) en ytterste visceral peritoneum eller serøs pels er laget av mesoteliale celler og tynt lag av bindevev. Det mangler i spiserøret,
ANNONSER:
(ii) Under dette er et muskulært lag dannet av glatte muskelfibre anordnet i ytre langsgående og indre sirkulære muskelfibre. Mellom de to lagene av muskler er et nettverk av nerveceller og nervefibre i det autonome nervesystemet, kjent som myenterisk plexus Eller plexus Av Auerbach.
(iii) Under muskellaget er en submukosa laget av bindevev med elastiske fibre, fett, blod og lymfekar, nerveceller og fibrekjertler,
(iv) Det innerste laget er en slimhinne som består av tre regioner:
(a) Ytterst smal muscularis mucosa av ytre langsgående og indre sirkulære glatte muskelfibre.
(B) Midtre tynt lag av lamina propria av bindevev, blodkar, nerver og knuter av lymfatisk vev, og
(c) en basalmembran som støtter et lag av søyle epitelceller som ofte er kjertel og ciliated.
Munn:
Munn er åpningen som fører inn i bukkalhulen. I lampreys (cyclostomes) er det en sirkulær åpning ved bunnen av bukkaltrakten og forblir permanent åpen på grunn av mangel på kjever, etc. I gnathostomes er det terminal. Munnen er avgrenset av lepper som er faste og dannet av cornified hud i fisk, amfibier og reptiler. Hos pattedyr er disse kjøttfulle og muskuløse.
Bukkal Hulrom:
ANNONSER:
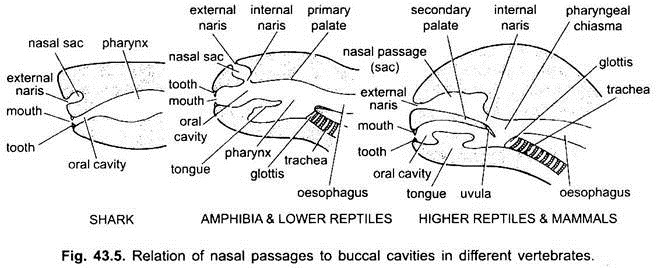
plassen mellom leppene og kjeftene er en vestibule. Det kan være avgrenset på utsiden av kinnene og på innsiden av tannkjøttet. Slimete kjertler i kinnene åpner inn i vestibulen. Munnen åpnes i et bukkal hulrom, som er et mellomrom mellom munnen og svelget. Det nøyaktige punktet hvor stomodeale ektoderm og pharyngeal endoderm fusjonerer er variabel og ikke lett å skille.
i elasmobranchs og de fleste benfisk åpner nesehulene ikke inn i bukkalhulen. I Chondrichthyes og tetrapoda (amfibier og de fleste reptiler) åpner nesehulene inn i bukkalhulen ved choanae eller indre narer, som er primitivt plassert forfra, men i krokodiller, fugler og pattedyr blir de bakre i svelget på grunn av dannelsen av en sekundær gane, som effektivt skiller respiratorisk nesepassasje fra munnhulen eller matpassasjen.
hos fugler er denne ganen spalt på grunn av hvilken nese-og bukkalhulrom kommuniserer med hverandre. I pattedyr fortsetter sekundær gane bakover som en membranøs myk gane. Hos mennesker henger myk gane inn i larynx-strupehodet i form av kjøttfull prosess, kalt drøvel.
Derivater Av Bukkal Hulrom:
1. Orale Kjertler:
det er to typer integumetary multicellular kjertler som åpner inn i bukkalhulen. De er slimete kjertler og enzymatiske kjertler. Fisk og akvatiske amfibier har bare slimete kjertler. Reptiler har kjertler i grupper, som palatine, lingual, sublingual og labial kjertler oppkalt etter plassering, de produserer også slim.
i giftige slanger er de øvre labialkjertlene modifisert for å utskille gift, mens I Gila-monsteret produserer sublinguale kjertler gift. Fugler har sublinguale kjertler og en kjertel i vinkelen av munnen. Pattedyr har mange små slimete kjertler i tillegg til ekte og forstørrede spyttkjertler som er enzymatiske. De er parotid, sublingual, submaxillary og infraorbital spyttkjertler, utskiller mucin og ptyalin.
2. Tunge:
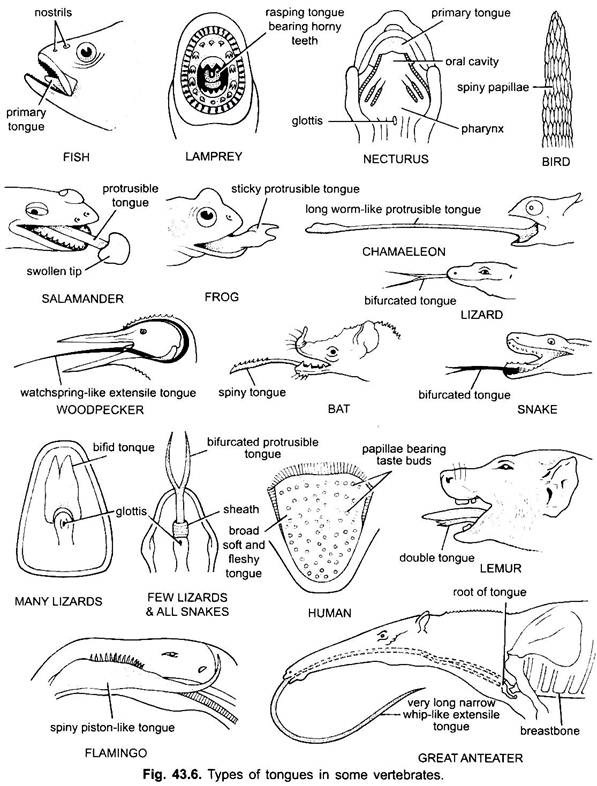
tungen finnes hovedsakelig i alle virveldyr. Tungen hos vertebrater viser mye mangfold og er ikke homologe. I syklostomer er det en muskuløs, kjøttfull, raspende tunge med kåte tenner for å rive huden og musklene i byttet.
Fisk har en primær tunge dannet av en kjøttfull fold av buccal gulvet. Det har ingen muskler, men reseptorer og tenner er tilstede på tungen i noen benfisk. Tungen er dekket med slimhinne. I noen amfibier er tungen enten mangelfull eller ubøyelig. De fleste amfibier har imidlertid en utstikkelig tunge, og i noen frosker og padder kan den foldes tilbake på seg selv når den ikke er i bruk.
det kan kastes ut av munnen ved rask tilførsel av lymf for å fange insekter. Tungen i øgler og slanger er ofte høyt utviklet. I kameleoner er det veldig utvidbart brukt til å fange insekter. Spissen er tykk og klebrig. Den forkede spissen av tungen i slanger tjener som et middel til å overføre kjemiske stimuli fra det ytre miljø til de parrede vomero-nasale organene på munntaket.
hos skilpadder og krokodiller kan ikke tungen forlenges. Amniote tungen har frivillige muskler, den mottar hypoglossal nerve og har kjertler og smaksløker. Det utvikler også indre muskler som beveger tungen. På fugler er tungen slank og har en kåt dekning. I noen fugler er tungen immobile, mens i noen fugler er den lang, protractil og ofte brukt til å fange maten.
i de fleste pattedyr, unntatt hvaler, er tungen høyt utviklet og i stand til betydelig bevegelse, i tillegg til forlengelse og tilbaketrekning, på grunn av tilstedeværelsen av en rekke indre muskler.
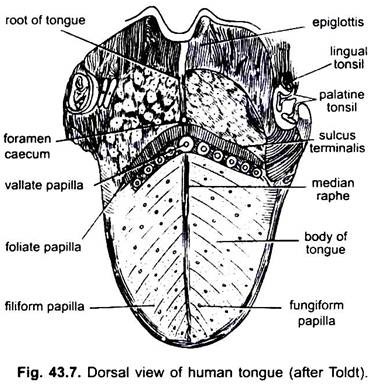
i pattedyr slimhinnen under tungen danner en median fold, kalt frenulum som knytter tungen til gulvet i munnen. I pattedyr bærer den øvre overflaten av tungen fire typer papiller, (filiform, fungiform, foliate og circumvallate), som bærer smaksløk unntatt filiform papiller.
3. Tenner:
Virveldyr har to typer tenner festet til kjevebein – epidermale tenner og ekte tenner. Epidermale tenner er best utviklet i syklostomer. De er harde, koniske, kåte strukturer avledet fra stratum corneum. I lampreys finnes de på bukkaltraktens vegger og på tungen. Tadpole larver av frosker og padder har serrated epidermale tenner i rader på leppene. Hos pattedyr har den voksne duckbill platypus epidermale tenner.
Ekte Tenner:
Tenner finnes ikke hos bardehval og anteaters hos pattedyr, og agnathans, steiner, noen padder, sirener, skilpadder og moderne fugler, etc. I lavere virveldyr (som fisk, amfibier og de fleste reptiler) tenner kan erstattes kontinuerlig et ubestemt antall ganger, slike tenner kalles polyphyodont. Disse tennene er homodont (lignende type) og acrodont. (med kjevebenene).
hos de fleste pattedyr er tennene diphyodont, thecodont og heterodont. I noen pattedyr er disse monofyodont å ha bare ett sett med tenner, f. eks mol, Indisk ekorn. Tennene er like i struktur til placoidskalaene av haier, dannet av et sentralt massehulrom, rundt hvilket er tilstede et tykt, men mykt lag, dentin, som er eksternt dekket av en tynn, ekstremt hard emalje. Disse er ment å ha avledet fra benete skalaer av ostracoderms og placoderms. For detaljer lesere kan se Tannsett I Pattedyr.
4. Adenohypophysis:
den fremre hypofysen utvikler seg som en dorsal evaginering av stomodaeum, kalt Rathkes pose, som innsnevrer seg for å danne de fremre og midtre lobene av hypofysen (adenohypophysis). Den bakre lobe av hypofysen eller neurohypophysis er ventral evaginering av diencephalon, kalt infundibulum. Dermed er det nervøs del.
Svelg:
delen av fordøyelseskanalen rett bak bukkalhulen er en svelg, foret med endoderm. Det er en vanlig passasje som tjener både for fordøyelsen og respirasjonen. Som en del av fordøyelsessystemet brukes den som en passasje for mat fra bukkalhulen til spiserøret, musklene begynner å svelge.
i fisk er svelget stort og lateralt perforert for gjellespalter, mens det i tetrapoda er kort og har åpninger i neseborene. I embryoer avgir svelgveggen en rekke evaginations som utvikler seg til spirakler, gill-clefts, luftblærer, lunger, mandler og noen endokrine kjertler (f.eks. thymus, skjoldbrusk og parathyroids).
Øsofagus:
spiserøret er kort hos de fleste fisk og amfibier fordi de mangler nakke, men det er lengre i amnioter på grunn av tilstedeværelse av nakke. Øsofagus av reptiler er lengre enn fisk og amfibier. I granivorøse og kjøttetende fugler forstørres en del av spiserøret i en sac-lignende pose kalt crop som tjener til å lagre mat som har blitt spist raskt.
avlingen mangler i hovedsak fordøyelseskjertler, selv om duene har 2 avlinger i begge kjønn, de er egentlig ikke kjertler, men celleformende strukturer, cellene danner ‘duemelk’ som blir matet til de unge. I pattedyr er spiserøret langt, mangler kjertler og varierer i forhold til lengden på nakken.
den passerer gjennom membranen, delen under membranen er dekket med visceral peritoneum som mangler fra den øvre delen. Øsofagus har slimete kjertler. Foringen danner langsgående folder, eller fingerlignende kjøttfulle papiller (elasmobranchs) eller kåt papiller i marine skilpadder.
Histologisk er øsofagus forskjellig fra resten av fordøyelseskanalen i tre fakta:
(i) det har ingen visceral peritoneum fordi det ligger utenfor coelom, dets ytterste dekklag er en tynn tunica adventitia.
(ii) muskelfibrene i den fremre delen er stripete, midtdelen har både stripete og ustripede muskler, og den bakre delen har bare ustripede muskler. Men det er unntak i drøvtyggere, alle musklene er stripete eller frivillige.
(iii) slimhinnen er laget av stratifiserte plateepitelceller og ikke av kolonneceller.
Mage:
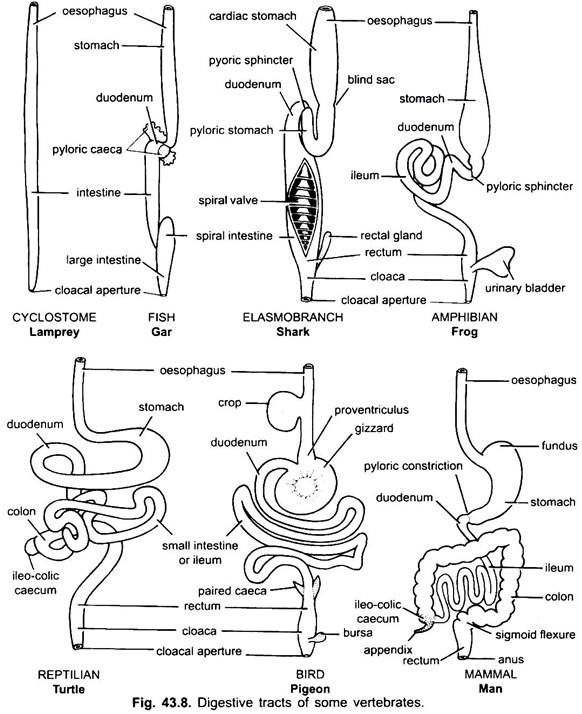
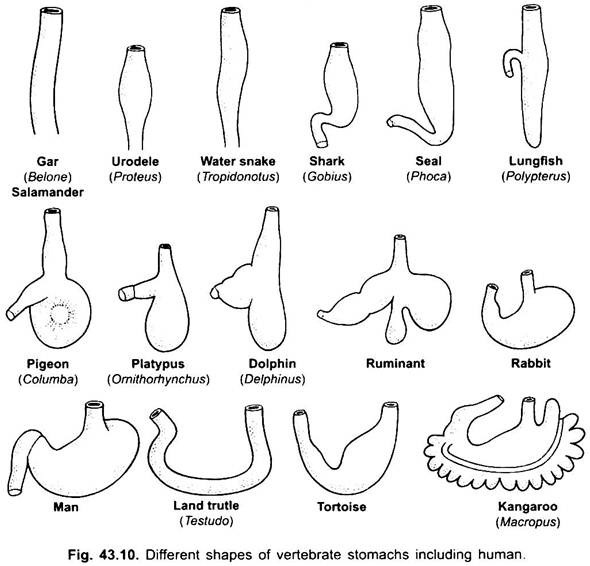
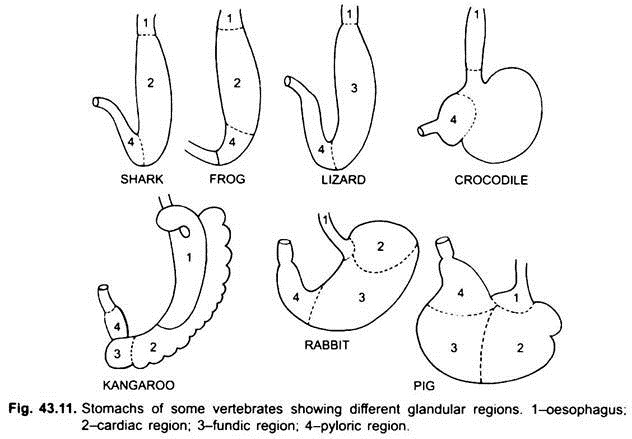
det er praktisk talt ingen mage i syklostomer, kimærer, lungefisk og noen primitive teleostfisk, siden den ikke har magekjertler, men i de fleste fisk og tetrapoda er den utvidet for lagring og macerasjon av fast mat og fordøyelse av mat fordi den inneholder magekjertler.
den første delen av magen, ved siden av spiserøret, er hjerteområdet og den nedre enden nær tarmen er pylorisk region, som har en pylorus eller pylorisk ventil hvor slimhinnen er omgitt av en tykk sphincter muskel som regulerer åpning og lukking av pylorisk mage i tarmen.
Magen er rett i syklostomer, gar, Belone, etc., og spindelformet I Proteus, Nekturus, noen øgler og slanger. I skilpadder og skilpadder er det et bredt buet rør, og i elasmobranchs er magen J-formet. I krokodiller og fugler har magen to deler, en proventriculus med magekjertler, og en svært muskuløs gizzard, som representerer pylorisk region og har en hard, cornified foring for sliping av mat.
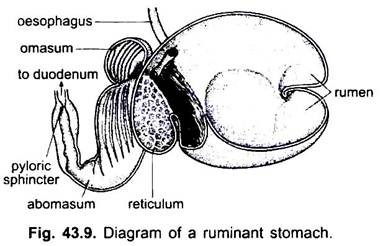
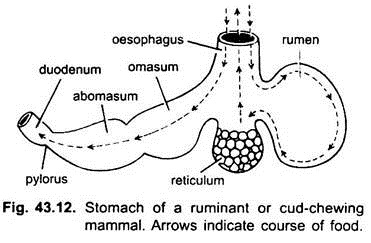
i pattedyr ligger magen på tvers og kan være en enkel sac eller delt inn i 3 regioner, nemlig hjerte -, fundisk og pylorisk, og hver region har sine magekjertler. I mange drøvtyggere har magen fire kamre – en rumen, retikulum, omasum og en abomasum. Det hevdes at de tre første kamrene er modifikasjoner av spiserøret, og abomasum er den sanne magen som representerer hjerte -, fundiske og pyloriske deler av magen.
det har blitt vist embryologisk at alle fire kamre er modifiserte regioner i magen. I kameler er det ingen omasum, vommen og retikulum har poselignende vannceller som en gang ble antatt å lagre vann, men de er sannsynligvis fordøyelses.
Histologisk har magen de typiske delene av fordøyelseskanalen, men den har to særegenheter, muscularis slimhinnen er laget av et ytre langsgående lag og et indre sirkulært lag av muskler. Epitelforingen er tykk med flere typer glandulære celler som danner magekjertler av tre typer kalt hjerte -, fundiske og pyloriske magekjertler.
hjerte-og pyloriske kjertler utskiller bare slim fra overflatecellene. Fundiske kjertler (eller hjertekjertler i noen) har tre typer celler, slimete nakkeceller produserer slim, oksyntiske celler produserer saltsyre, de kan være tilstede i hjerteområdet også, zymogenceller eller peptiske celler utskiller pepsin.
hos de fleste dyr utskiller zymogencellene også to proenzymer kalt propepsin og prorennin som omdannes av saltsyre til henholdsvis pepsin og rennin. Sekresjonene av alle mageceller danner en blanding som kalles magesaft.
Tynntarm:
Tynntarmen er lang, smal og kveilet rør etter pylorus. Det er den viktigste delen av fordøyelseskanalen fordi fordøyelsen og absorpsjonen av mat finner sted i den. I syklostomer er tarmen et kort rett rør med en spiralformet langsgående klaff som strekker seg inn i den.
i elasmobranchs er den delt inn i små og store deler, og den lille delen har en spiralventil som øker den absorberende overflaten sterkt. En spiralventil er også tilstede i tynntarmen til noen få primitive benfisk, men mangler i høyere former hvor tarmen er lang og kveilet.
i caecilians er det lite kveilet og ikke differensiert til en liten og stor kanal. I frosker og padder er den relativt lang og kveilet. I reptiler er det mer spolet enn i amfibier. For første gang i vertebrater oppstår et caecum eller blind divertikulum ved krysset mellom små og store tarm.
dette er imidlertid ikke permanent i alle reptiler. Hos fugler er tynntarmen viklet eller sløyfet, og en eller to kolikk caeca er også tilstede ved krysset mellom små og store tarm. Hos de fleste pattedyr er også tynntarmen forholdsmessig lang og kveilet. Dens lengde er imidlertid korrelert med matvaner. I planteetere er det relativt lengre i forhold til insektsdyr og rovdyr.
det er en blind lomme eller cecum i krysset av tykktarmen og tynntarmen som er generelt liten i kjøttetende arter og ganske lang i mange planteetere. Den første delen av tynntarmen er tolvfingertarmen, som er kort begynnelse fra pylorus og slutter utenfor inngangen til bukspyttkjertel og leverkanaler.
Den har mange foldede villi og inneholder forgrenende Brunners kjertler i submukosa som utskiller slim, noe alkalisk vannaktig væske og et lite enzym. Tolvfingertarmen produserer også to hormoner kalt secretin og cholecystokinin som stimulerer bukspyttkjertelen og galleblæren til å frigjøre sine juice. Kanaler fra galleblæren og bukspyttkjertelen åpner inn i tolvfingertarmen.
bak tolvfingertarmen er en ileum, som bare hos pattedyr er differensiert til en fremre mindre jejunum og bakre lengre ileum. Et stort antall små fordøyelseskjertler er tilstede i tynntarmen. De er rørformede kjertler Eller krypter Av Lieberkuhn funnet gjennom hele lengden, de utskiller slim og en succus entericus som har flere enzymer.
foringen av tynntarmen foldes for å danne små villi, noe som øker overflaten for sekresjon og absorpsjon. Villi er dekket tett av minutt finger-lignende anslag, kalt microvilli som bistå i absorpsjon i villi. I pattedyr finnes knuter av lymfoid vev kalt Peyers flekker på ileum.
Tykktarmen:
Tykktarmen har en større diameter enn tynntarmen. Det er generelt kort i fisk, amfibier, reptiler og fugler, men i pattedyr er det langt. I nedre former danner tykktarmen en endetarm, men i tetrapoda har den en kolon og terminal endetarm. I de fleste fisk og amfibier fører den terminale delen av endetarmen til en cloaca dannet av proctodaeum.
rektum, ekskretjonskanaler og kjønnskanaler åpner seg inn i cloaca, og den åpner til utsiden med en cloacal åpning. Men i mange benfisk og alle pattedyr (unntatt monotremes) har rektum og urinogenitale kanaler separate åpninger til utsiden; åpningen av den tidligere er en anus.
Rektum av pattedyr er ikke homolog med rektum av vertebrater siden i pattedyr er det avledet ved partisjonering av embryonale cloaca. I de fleste vertebrate embryoer er det en postanal tarm som en forlengelse av tarmen i halen, men den forsvinner senere.
i elasmobranchs bærer tykktarmen et par rektale kjertler som utskiller slim og natriumklorid. I amnioter er det en ileokolisk ventil ved krysset mellom små og store tarm, som er fraværende i fisk. Det forhindrer bakterier å komme inn i ileum fra kolon.
i amniotes fra dette krysset oppstår en ileocolic caecum som er to i fugler. Den inneholder cellulose fordøye bakterier. Det er veldig lenge i plantelevende pattedyr (kanin, hest, ku, etc.). I primater er caecum liten med et vestigial vermiform vedlegg.
Fordøyelseskjertler:
1. Lever:
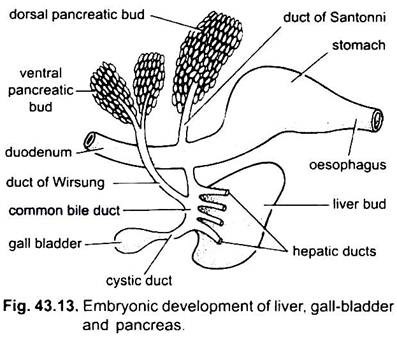
leveren oppstår som en enkelt eller dobbel utvekst fra den ventrale veggen av den embryonale archenteron. Denne utveksten danner et hul hepatisk divertikulum, som snart skiller seg ut i en fremre del, som prolifererer til å bli leveren og dens gallekanaler, og en bakre del som gir opphav til galleblæren og cystisk kanal. Gallegangene blir med for å danne en leverkanal som forener med cystisk kanal for å danne en felles gallekanal eller ductus choledochus. Regionen av archenteron hvorfra leveren oppstår, blir tolvfingertarmen.
leveren er den største lobed kjertelen i kroppen, suspendert av et dobbeltlag av peritoneum fra tverrgående septum eller dets representant.
en galleblære er for lagring av galle utskilt av leverceller, ligger i leveren og drenerer inn i tolvfingertarmen gjennom felles gallekanal dannet ved forening av cystisk kanal og leverkanal. En galleblære er ikke uunnværlig og mangler i mange fugler og pattedyr.
en lever er tilstede hos alle vertebrater. I cyclostomer er det lite, enkelt flikete (lampreys) og to flikete i hagfishes. Det er bilobed i elasmobranchs, to eller tre flikete i bony fisk, amfibier, reptiler og fugler og mange flikete i pattedyr. Leveren er lang, smal og sylindrisk i fisk, urodeles og slanger.
Den er kort, bred og flat i fugler og pattedyr. En galleblære og gallekanal er tilstede i larvcyklostomer, men de er fraværende hos voksne. Fisk, amfibier og reptiler har generelt en galleblære, men det mangler i mange fugler. De fleste pattedyr har en galleblære, men det er fraværende I Cetacea og Ungulata.
leveren utskiller en vannaktig, alkalisk galle, men har ingen enzymer. Det nøytraliserer surheten av mat som kommer inn i tolvfingertarmen. Det hjelpemidler i fordøyelsen av fett.
2. Bukspyttkjertel:
Bukspyttkjertel er dannet fra endodermen til den embryonale archenteron. Et enkelt dorsalt divertikulum fra embryonalt tolvfingertarm og en eller to ventrale utvekster fra leveren danner bukspyttkjerteldivertikula. De proksimale delene av divertikula danner bukspyttkjertelkanaler, men disse kanalene gjennomgår reduksjon eller fusjon, slik at bare en eller to bukspyttkjertelkanaler forblir i voksen, de åpner i tolvfingertarmen enten separat eller etter forening med den vanlige gallekanalen.
de distale delene av divertikula gjennomgår spirende for å danne hovedmassen av bukspyttkjertelceller som mesodermale derivater tilsettes. Således er en enkelt kjertel laget som har flere lober som danner enten en diffus eller en kompakt bukspyttkjertel.
bukspyttkjertelen er både en eksokrin og endokrin kjertel, bundet sammen av delikate tråder av bindevev. Den eksokrine delen utskiller fordøyelsesenzymer som helles i tolvfingertarmen gjennom bukspyttkjertelkanaler. Mens den endokrine delen utskiller hormoner som insulin og glukagon.
en bukspyttkjertel er tilstede hos alle vertebrater. I lampreys, noen benfisk, lungfisk og lavere tetrapoder, er det et diffust organ innebygd i leveren, mesenterier og tarmvegg. Hagfisk har en liten bukspyttkjertel. Elasmobranchs har en veldefinert bilobed bukspyttkjertel. I høyere tetrapoda er det generelt en kompakt kjertel. En eller to bukspyttkjertelkanaler åpner i tolvfingertarmen.