ANNONCES:
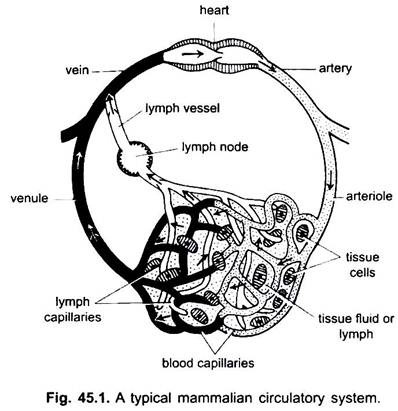
Dans le système circulatoire des vertébrés, il existe deux systèmes de tubes à ramifications complexes, qui se ramifient dans tout le corps et transportent les fluides vers les tissus. Ils sont un système vasculaire sanguin et un système lymphatique. Lorsque le terme système circulatoire est utilisé, il se réfère uniquement au système vasculaire sanguin.
Le système vasculaire sanguin est un système fermé chez les vertébrés; il a un cœur contractile et des tubes continus appelés vaisseaux. Le système lymphatique est un système ouvert avec des espaces lymphatiques. Les vaisseaux sanguins qui transportent le sang loin du cœur sont des artères, qui se divisent en artérioles plus minces, se ramifiant en capillaires extrêmement fins et petits.
La paroi d’un capillaire est constituée d’une seule couche de cellules endothéliales à glands. Chaque vaisseau sanguin, y compris le cœur, a une paroi de cellules endothéliales ou d’endothélium. Le sang n’entre pas directement en contact avec les cellules tissulaires.
ANNONCES:
Les substances passent de et dans les capillaires à travers le liquide tissulaire contenu dans les espaces tissulaires entre les cellules. L’échange de substances entre les cellules sanguines et tissulaires du corps a lieu à travers les parois capillaires. Ceci est provoqué par une pinocytose dans les cellules endothéliales des capillaires.
Au microscope électronique, les cellules endothéliales montrent de nombreuses petites vésicules qui sont des invaginations des membranes plasmiques. Ces vésicules se déplacent d’un côté de la cellule et libèrent leur contenu du côté opposé.
Ainsi, le sang donne de l’oxygène, du matériel nutritif et des hormones aux cellules pour le métabolisme, et les cellules donnent de l’eau, du dioxyde de carbone et des déchets azotés dans le sang pour les transporter vers les organes excréteurs pour leur élimination rapide.
Les capillaires forment un réseau dans tous les tissus corporels à l’exception du cartilage et de l’épithélium. À partir des capillaires, le sang passe en fines veinules qui se combinent pour former des veines qui transportent le sang vers le cœur. Mais certaines veines (veines portales, veines rénales et veine hépatique) ont des capillaires qui ressemblent à ceux des artères. Mais tout le sang ne passe pas à travers les capillaires dans les veinules.
ANNONCES:
Il existe également des canaux traversants entre les artérioles et les veinules situés dans certains organes, tels que la peau. Il existe également des anastomoses artério-veineuses entre les artérioles et les veinules trouvées dans les chiffres. La fonction de telles connexions n’est pas clairement connue, bien qu’il soit affirmé qu’elles régulent la pression artérielle et la circulation.
À certains endroits, l’échange de matériel entre les tissus et le sang a lieu à travers des espaces à parois minces ou des sinusoïdes. Dans certains organes, un vaisseau sanguin peut former un réseau enroulé de minuscules vaisseaux sanguins appelés rete mirabile (reins, vessie).
Parties du Système circulatoire:
ANNONCES:
Chez les chordés, le système circulatoire est de type fermé. Ce type se trouve également chez les annélides (invertébrés). Un autre type est de type ouvert dans lequel les capillaires ne se trouvent pas. On le trouve chez les mollusques et les arthropodes. Ici, le sang circule à travers les artères dans divers organes, qui traversent des espaces sanguins ou des sinus, puis à nouveau dans des vaisseaux (veines) jusqu’au cœur.
Le système circulatoire comprend le cœur, les artères, les veines, les capillaires et le sang. Le cœur est un vaisseau sanguin modifié avec des parois musculaires, qui se contracte périodiquement pour pomper le sang dans les différentes parties du corps à travers des vaisseaux définis.
Les artères et leurs branches forment un système artériel qui transporte le sang du cœur. Les veines et leurs affluents constituent un système veineux qui prélève le sang des capillaires des artères ou des artérioles et le transporte jusqu’au cœur.
Système portal:
Dans le système portal, le sang n’est pas renvoyé directement vers le cœur, mais il y a un organe interposant (foie ou rein) au cours du retour du sang. La veine apportant le sang commence dans les capillaires et se termine dans les capillaires, la veine concernée agissant à la fois comme vaisseau afférent et efférent, les vaisseaux afférents se terminent dans les capillaires tout comme les artères, puis le sang est recueilli dans les veines systémiques.
Tous les vertébrés ont un système portail hépatique dans lequel le sang passe dans deux ensembles de capillaires dans le foie. Les vertébrés inférieurs et les embryons de vertébrés supérieurs ont également un système de portail rénal dans lequel le sang traverse deux ensembles de capillaires dans le rein avant d’atteindre le cœur. Les capillaires de l’hypophyse forment un système de portail hypophysaire qui est un système petit mais important.
Système lymphatique :
On le trouve chez les chordés à l’exception des cyclostomes et des poissons cartilagineux. Il comprend la lymphe et les vaisseaux lymphatiques. La lymphe est un liquide tissulaire présent parmi les cellules du corps. C’est le plasma sanguin moins les globules rouges et certaines protéines.
Capillaires lymphatiques formant un réseau de minces vaisseaux aveugles qui recueillent la lymphe. Les vaisseaux lymphatiques sont des vaisseaux à paroi mince formés par l’union de capillaires sanguins. Ceux-ci se vident dans les veines. Les ganglions lymphatiques se trouvent sur les vaisseaux lymphatiques chez les mammifères. Ceux-ci forment des lymphocytes du sang utilisés pour la défense du corps contre les maladies.
Évolution du Cœur chez les vertébrés:
ANNONCES:
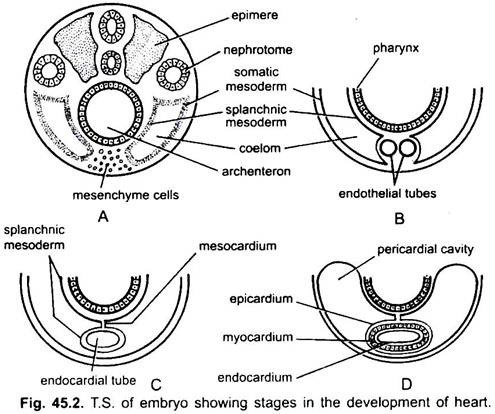
Le cœur est un organe non apparié mais son origine est bilatérale. Dans un embryon, le mésenchyme forme un groupe de cellules endocardiques sous le pharynx. Ces cellules s’arrangent pour former une paire de tubes endothéliaux minces. Les deux tubes endothéliaux fusionnent rapidement pour former un seul tube endocardique situé longitudinalement sous le pharynx.
Le mésoderme splanchnique situé sous l’endoderme est replié longitudinalement autour du tube endocardique. Ce tube à deux couches formera le cœur dans lequel le mésoderme splanchnique s’épaissit pour former un myocarde ou une paroi musculaire du cœur et un épicarde mince externe ou un péricarde viscéral. Le tube endocardique devient la muqueuse du cœur appelée endocarde.
Les plis du mésoderme splanchnique se rencontrent au-dessus pour former un mésocarde dorsal qui suspend le cœur dans le cœlome. Bientôt, un septum transversal se forme derrière le cœur qui divise le cœlome en deux chambres, une cavité péricardique antérieure enfermant le cœur et une cavité abdominale postérieure.
Le cœur est un tube droit mais il augmente en longueur et devient en forme de S car ses extrémités sont fixes. L’apparition de valves, la constriction, les cloisons dans le cœur et les épaississements différentiels de ses parois forment trois ou quatre chambres dans le cœur.
1. Cœur à chambre unique:
Chez Amphioxus (chordé primitif), on ne trouve pas de véritable cœur. Une partie de l’aorte ventrale sous le pharynx est musculaire et contractile et agit comme cœur.
2. Coeur À Deux Chambres:
Dans les cyclostomes, il y a quatre chambres disposées dans un ordre linéaire – un sinus venin à paroi mince, un oreillette légèrement musculaire (oreillette), un ventricule musculaire et un conus artériel musculaire ou bulbus cordis. Il se trouve dans la cavité corporelle dans laquelle d’autres organes viscéraux sont également présents.
Sur quatre chambres, seuls l’oreillette et le ventricule correspondent aux quatre chambres (oreillettes appariées et ventricules appariés) des vertébrés supérieurs. Dans l’évolution du cœur, de nombreux changements ont eu lieu.
Élasmobranches:
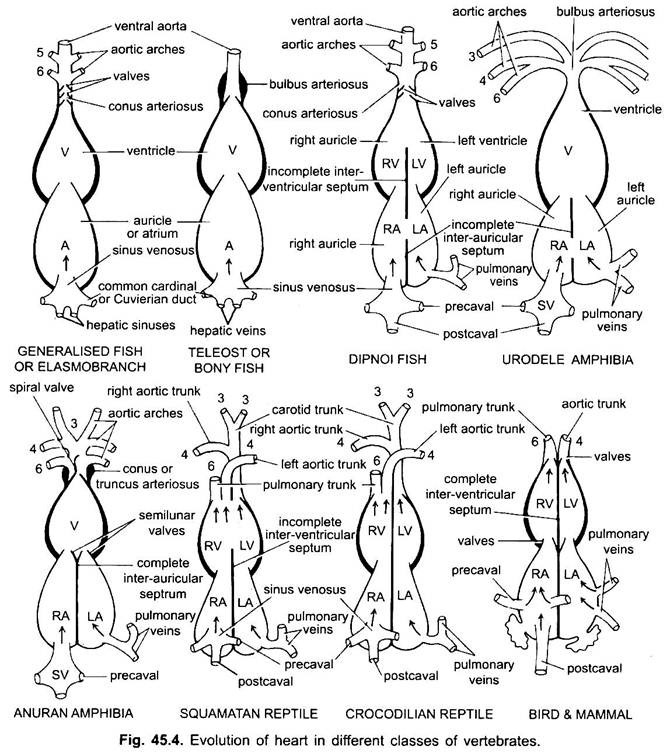
À l’exception des Dipnoi, le système circulatoire des poissons des cyclostomes aux téléostéens, seul le sang non oxygéné va au cœur, de là il est pompé vers les branchies, aéré puis distribué au corps. Le cœur de l’aiguillat cartilagineux est un tube en forme de S musclé et courbé dorsoventralement avec quatre compartiments en série linéaire.
Ce sont le sinus venin et l’oreillette pour recevoir le sang veineux, et un ventricule et un conus artériel pour pomper ce sang. Le cœur est un cœur veineux branchial. Le sinus venosus et le conus artériel sont des chambres accessoires. L’oreillette et le ventricule sont de véritables chambres, c’est donc un cœur à 2 chambres.
Le sinus venin s’ouvre antérieurement dans l’oreillette par une ouverture sinus-auriculaire gardée par une paire de valves. L’oreillette est dorsale au ventricule et s’ouvre ventralement dans le ventricule par une ouverture auriculo-ventriculaire gardée par une paire de valves. Le ventricule musculaire à paroi épaisse s’ouvre dans un étroit conus artériel contenant des valves en deux séries.
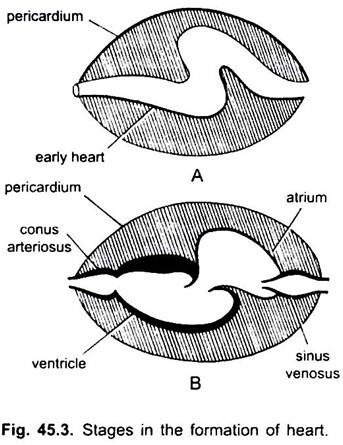
Le cœur est enfermé dans une cavité péricardique séparée de la cavité corporelle par un septum transversal. Conus perce le péricarde et devient continu avec l’aorte ventrale. La cavité péricardique communique avec la cavité corporelle par deux perforations dans le septum transversal.
Téléostéens :
Leur cœur ressemble à celui des clasmobranches. Dans les téléostéens, le conus est réduit et a une seule paire de valves. La partie proximale de l’aorte ventrale près du conus devient fortement élargie et à paroi épaisse, appelée bulbus artériel. Il est élastique et se dilate au moment de la contraction ventriculaire. Le cœur est donc à 2 chambres avec une seule circulation de sang.
3. Coeur à trois chambres:
Dipnoi:
Chez les diphoanes, un septum divise l’oreillette en une chambre droite et une chambre gauche. Ceci est corrélé à l’utilisation de la vessie natatoire comme organe de respiration et représente la première étape vers le développement du système circulatoire à double type, dans lequel le sang oxygéné et non oxygéné pénètre dans le cœur et est maintenu séparé.
Le sang de l’oreillette droite du poisson-poumon passe dans le ventricule droit et est ensuite pompé dans la vessie gazeuse primitive ressemblant à un poumon par les artères pulmonaires qui se ramifient à partir de la sixième paire d’arcs aortiques. Le sang oxygéné retourne dans l’oreillette gauche par le biais de veines pulmonaires comme les amphibiens.
Amphibies:
Chez les amphibiens, l’oreillette dorsale se déplace en avant du ventricule. Le sinus venosus s’ouvre dans l’oreillette droite dorsalement et non postérieurement. L’oreillette est complètement divisée en chambres droite et gauche et n’a pas de foramen ovale dans le septum inter-auriculaire, qui reste ouvert chez les dipnoaires.
Des poches profondes se développent dans la cavité ventriculaire. Le conus artériel se divise en vaisseaux systémiques et pulmonaires par une valve en spirale. Chez les salamandres moins pulmonaires, le septum inter-auriculaire est incomplet et les veines pulmonaires sont absentes.
Reptilia:
Chez les reptiles, le cœur est plus avancé. L’oreillette est toujours complètement séparée en une chambre droite et une chambre gauche, et sous de nombreuses formes, le sinus veneux est incorporé dans la paroi de l’oreillette droite. Le ventricule est également partiellement divisé par un septum chez la plupart des reptiles, et chez les alligators et les crocodiles est complètement à deux chambres.
Cela signifie que le sang oxygéné provenant des poumons vers le côté gauche du cœur est essentiellement séparé du sang non oxygéné du corps vers le côté droit. Ainsi, chez les crocodiliens, les deux types de sang sont complètement séparés et presque complets chez les autres reptiles, mais un certain mélange se produit dans d’autres parties du système circulatoire.
Le conus artériel embryonnaire se divise en trois vaisseaux au lieu de deux:
(i) Arcade pulmonaire transportant le sang vers les poumons du côté droit du ventricule.
(ii) Aorte systémique droite transportant le sang du côté gauche du ventricule vers le corps par le quatrième arc aortique droit.
(iii) Le système systémique gauche provient du ventricule droit vers le quatrième arc aortique gauche.
Au point de contact avec l’aorte systémique du ventricule gauche, même chez les crocodiliens, une ouverture entre les deux est présente, appelée foramen des Panizzae où il peut y avoir un mélange des deux types de sang. Ainsi, le cœur reptilien représente le cœur de transition contre le cœur amphibien – 2 oreillettes complètes et 2 ventricules incomplets avec un peu de mélange de sang à droite et à gauche systémique.
4. Cœur à quatre chambres:
Aves et Mammalia:
Chez les oiseaux, le ventricule est complètement divisé en deux, de sorte que le cœur est à quatre chambres (2 oreillettes et 2 ventricules). Il y a une séparation complète du sang veineux et artériel. L’aorte systémique quitte le ventricule gauche et transporte le sang vers la tête et le corps. Alors que l’artère pulmonaire quitte le ventricule droit et transporte le sang vers les poumons pour l’oxygénation.
Ainsi, il y a une double circulation dans laquelle il n’y a aucun mélange de sang à aucun endroit. Le sinus venin est complètement incorporé dans l’oreillette droite, qui reçoit deux précavales et une postcavale. L’oreillette gauche reçoit du sang oxygéné par les veines pulmonaires, le conus artériel est absent, l’aorte pulmonaire provient du ventricule droit et une aorte systémique unique provient du ventricule gauche et les deux ont des valves à leur base.
Modifications des arcs aortiques chez les vertébrés:
Artères embryonnaires:
Lors de la formation du cœur chez un embryon de vertébré, un vaisseau sanguin appelé aorte ventrale apparaît à mi-ventrale sous le pharynx, qui se connecte rapidement au conus artériel. L’aorte ventrale provenant du cœur s’avance sous le pharynx et se divise antérieurement en une paire d’artères carotides externes dans la tête.
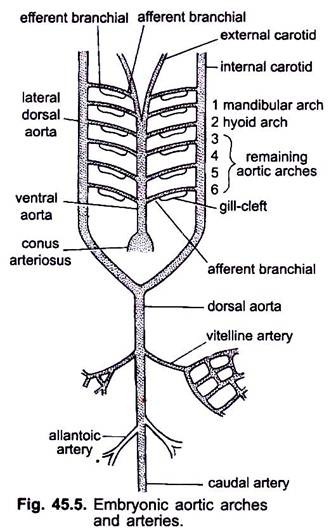
L’aorte ventrale dégage 6 paires d’arcades aortiques latérales à équidistance qui traversent les arcades viscérales. Chaque arc aortique a une artère branchiale afférente ventrale transportant le sang veineux vers la branchie et une artère branchiale efférente dorsale prélevant le sang oxygéné de la branchie. Les artères branchiales efférentes de chaque côté se rejoignent dorsalement avec l’aorte dorsale latérale ou l’aorte radix qui pénètre dans la tête en tant qu’artère carotide interne.
Le premier arc aortique est un arc aortique mandibulaire, le second est un arc aortique hyoïde, les autres sont appelés troisième, quatrième, cinquième et sixième arcs aortiques. L’aorte dorsale latérale fusionne derrière le pharynx pour former une aorte dorsale qui se poursuit à mi-dorsalement dans la queue en tant qu’artère caudale.
De l’aorte dorsale apparaissent des artères appariées et non appariées qui alimentent divers organes du corps. Dans un embryon avec un sac vitellin, une paire d’artères vitellines naît de l’aorte dorsale et alimente le sac vitellin. Chez les embryons d’amniotes, une paire d’artères ombilicales ou allantoïques provient de l’aorte dorsale fournissant du sang à l’allantois.
Chez un adulte, les artères vitellines fusionnent pour former l’artère mésentérique principale, la majeure partie des artères allantoïques est perdue, mais leurs restes forment des artères iliaques hypogastriques ou internes.
Arcs aortiques chez les vertébrés:
Chez divers vertébrés adultes, le système artériel semble différent, mais ils sont construits sur le même plan fondamental de base. La différence est due à la complexité croissante du cœur due à un changement de la respiration des branchies aux poumons. Il y a une réduction progressive du nombre d’arcs aortiques dans la série des vertébrés.
Cyclostomata:
Chez Petromyzon, il y a 7 paires d’arcs aortiques. Dans d’autres cyclostomes, ceux-ci varient de 6 paires chez la Myxine et de 15 paires chez Eptatretus.
Poissons:
Bien que six soit considéré comme le nombre de base d’arcs aortiques pour les poissons. Ce nombre est réduit à cinq même chez les requins et les raies avec la perte de la première paire, l’arc aortique mandibulaire ou il est représenté par une artère pseudobranchiale efférente. Chez la plupart des poissons osseux, les arcs aortiques mandibulaires (i) et hyoïdes (ii) disparaissent ou sont très réduits.
Chez les polyptères et les Dipnoi (poissons pulmonaires), les branchies ne sont pas bien développées. Ainsi, l’artère pulmonaire provient de la partie efférente du 6ème arc de chaque côté qui fournit du sang à la vessie ou au poumon. Chez les élasmobranches et les Dipnoi, chaque arcade a une artère branchiale afférente et deux artères branchiales efférentes (formées par division) dans chaque branchie. Chez les poissons osseux, chaque branchie a une artère afférente et une artère efférente.
Chez les tétrapodes, les arcades aortiques ne se divisent pas en parties afférentes et efférentes car les véritables branchies internes sont absentes. Dans tous les tétrapodes, les premier et deuxième arcs disparaissent.
Amphibies:
Ici, les arcs aortiques montrent une modification due à la perte des branchies et à l’apparence des poumons. Chez les urodèles, des branchies externes sont présentes en tant qu’organes respiratoires en plus des poumons. Les arcs aortiques III, IV, V et VI sont présents, bien que la cinquième paire soit très réduite chez Siren, Amphiuma et Necturus. Les arcs aortiques ne sont pas brisés dans les branchies externes en parties afférentes et efférentes, car les branches issues des arcs aortiques IV, V et VI forment des capillaires dans les branchies externes.
Les aortes dorsales latérales entre les arcs aortiques III et IV persistent en tant que connexion vasculaire, le canal carotique. L’arc aortique VI forme l’arc pulmo-cutané ou l’artère de chaque côté qui transporte le sang vers les poumons et la peau. Il conserve également une connexion avec l’aorte dorsale latérale connue sous le nom de canal artériel (canal de Botalli).
Chez la larve d’anuran (têtard de grenouille), la disposition des arcs aortiques ressemble à une urodèle adulte en raison de la présence de branchies. Lors de la métamorphose, avec la perte des branchies, les arcs aortiques I, II et V disparaissent complètement, seuls les IIIème, IVème et Vlème arcs aortiques sont présents. L’aorte dorsale latérale entre les troisième et quatrième arcades aortiques (ductus caroticus) disparaît également. Ainsi, la troisième arcade aortique avec une partie de l’aorte ventrale devient l’arcade carotide transportant le sang oxygéné vers la région de la tête.
Le quatrième arc aortique le long de son aorte dorsale latérale forme l’arc systémique. Le sixième arc aortique devient l’arc pulmocutané fournissant du sang veineux aux poumons et à la peau. Le canal artériel disparaît lors de la métamorphose. Ainsi, les anoures adultes n’ont que des arcs aortiques III, IV et VI. Ceux-ci sont également retenus par les amniotes.
Reptilia:
Chez les reptiles, les branchies sont entièrement remplacées par des poumons. Seuls les arcs aortiques III, IV et VI sont présents. Avec la séparation partielle du ventricule en deux parties, la partie distale du conus artériel et l’ensemble de l’aorte ventrale sont divisés en trois vaisseaux, c’est-à-dire deux aortiques ou systémiques et un pulmonaire. L’arcade systémique droite (IV) provient du ventricule gauche transportant du sang oxygéné vers l’arcade carotide (III). L’aorte systémique gauche (IV) et l’aorte pulmonaire (VI) prennent leur origine du ventricule droit. Le système gauche transporte le sang désoxygéné ou mélangé vers le corps par l’aorte dorsale.
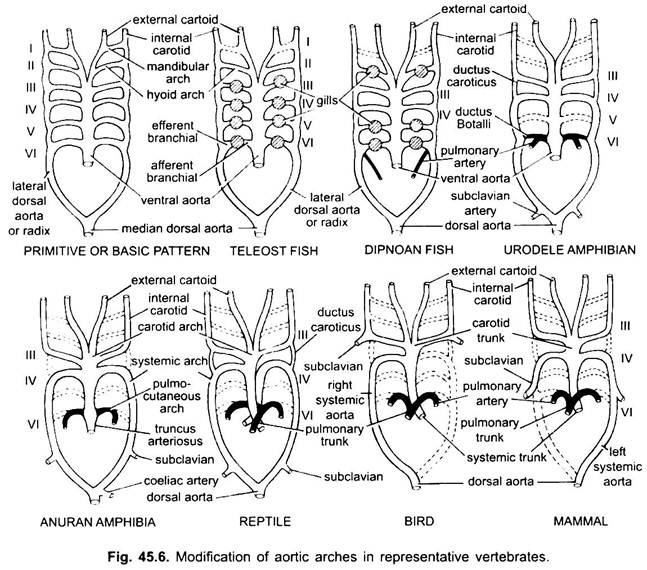
Pendant que l’artère pulmonaire transporte le sang désoxygéné vers les poumons. Le canal carotique disparaît, mais il persiste chez les serpents et certains lézards (Uromastix). Le canal artériel disparaît chez la plupart des reptiles bien qu’il persiste sous une forme réduite chez les sphénodons et certaines tortues. En raison du mélange de sang, les reptiles sont des animaux à sang froid comme les poissons et les amphibiens.
Aves:
Chez les oiseaux, les arcs aortiques III, IV et VI sont présents. Ils suivent le modèle général des reptiles avec quelques différences. Avec la division complète du ventricule en deux parties, le conus artériel et l’aorte ventrale se sont divisés pour former deux vaisseaux, l’aorte systémique provenant du ventricule gauche et une aorte pulmonaire provenant du ventricule droit.
Un troisième arc aortique avec des restes d’aorte latérale et ventrale forme les carotides issues de l’aorte systémique. La quatrième arche aortique forme l’aorte systémique du côté droit seulement. Il s’unit à l’aorte latérale de son propre côté et forme l’aorte dorsale. Une partie de la quatrième arche aortique du côté gauche forme l’artère sous-clavière gauche, le reste avec son aorte dorsale latérale disparaît. Le sixième arc aortique forme l’aorte pulmonaire. Le canal carotique et le canal artériel disparaissent.
Mammalia:
Chez les mammifères, les arcs aoriques III, IV et VI persistent également. Le ventricule est complètement divisé en deux parties.
Le conus artériel et l’aorte ventrale se séparent pour former deux vaisseaux:
(i) Une aorte systémique provenant du ventricule gauche et
(ii) Une aorte pulmonaire provenant du ventricule droit. La troisième arcade aortique avec des restes d’aorte latérale et ventrale forme l’arcade carotide.
La quatrième arche aortique forme l’aorte systémique du côté gauche seulement, tandis que du côté droit sa partie proximale forme une artère sous-clavière innominée et droite, le reste avec son aorte dorsale latérale disparaît. Le sixième arc aortique forme l’aorte pulmonaire. Le canal artériel dégénère mais il persiste chez certains jusqu’à l’éclosion ou la naissance sous une forme réduite sur le côté gauche sous la forme d’un ligament artériel mince.
Système veineux:
Veines embryonnaires:
Chez tous les embryons de vertébrés, le système veineux est simple et similaire. Les veines sont pour la plupart appariées et disposées symétriquement. Chez les embryons sans sac vitellin, une veine sous-intestinale se forme dans le mésoderme splanchnique sous l’intestin.
Il boucle autour de l’anus et se poursuit postérieurement comme:
(i) Une veine caudale dans la queue. Chez tous les embryons ayant un sac vitellin (contenant ou non du jaune), une paire de veines vitellines naît du sac vitellin et rejoint la partie postérieure du cœur en développement qui devient le sinus venin. En fait, la fusion de ces veines vitellines est responsable de la formation du cœur chez les poissons osseux, les reptiles et les oiseaux. Chaque veine vitelline à son extrémité postérieure rejoint la veine sous-intestinale formée de la même manière que dans les embryons sans sac vitellin.
(ii) Une paire de veines subcardinales apparaît entre les reins et rejoint la veine caudale.
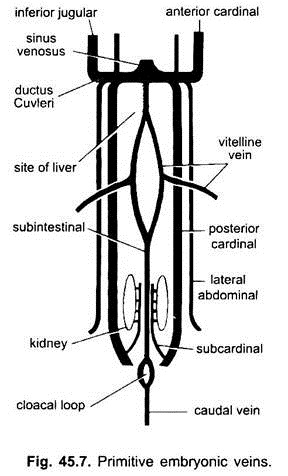
( iii) Des veines cardinales antérieures et postérieures appariées sont formées, qui transportent le sang de la tête et des parties postérieures du corps respectivement. Les veines cardinales antérieure et postérieure de chaque côté s’unissent pour former un canal Cuvieri ou veine cardinale commune qui passe vers l’intérieur à travers le septum transversal pour entrer dans le sinus venosus.
Chez les poissons et les salamandres (urodèles), une veine jugulaire inférieure vient du côté ventral de la tête pour rejoindre la veine cardinale commune. Il n’a pas d’homologue chez les autres vertébrés. Chez un amniote, une paire de veines abdominales latérales ou ventrales sort de la paroi du corps pour pénétrer dans les veines cardinales communes.
Poissons:
La veine cardinale commune (canal de Cuvier) pénètre dans le sinus veneux de chaque côté et est formée par la fusion des cardinaux antérieur et postérieur. Le sang de la tête est collecté par les cardinaux antérieurs, et le sang des reins et des gonades est collecté par les cardinaux postérieurs.
Les veines abdominales latérales appariées qui reçoivent le sang de la paroi du corps et des appendices appariés pénètrent également dans les canaux de Cuvier. Le système porte rénal est constitué de la veine caudale et de deux veines porte rénales situées latéralement aux reins qui se capillarisent dans les reins.
Le système portail hépatique transporte le sang de l’estomac et de l’intestin et le renvoie au foie, où, après avoir traversé des séries de sinusoïdes, il pénètre dans le sinus venin par des veines hapatiques appariées.
Chez les téléostéens, les veines abdominales latérales manquent de sorte que le sang des sous-clavières, drainant les appendices pectoraux, pénètre directement dans le sinus venosus et que le sang des veines iliaques, drainant les appendices pelviens, passe dans les postcardinaux.
Chez les dipnoaires, une seule veine abdominale ventrale est présente, vraisemblablement dérivée d’une fusion d’abdominaux latéraux. Cette veine reçoit le sang des ilacs par le biais de veines pelviennes appariées et pénètre dans le canal droit de Cuvier. De plus, à partir du système postcardinal droit, une nouvelle veine, le postclaval fait son apparition qui est d’une importance majeure chez les vertébrés supérieurs. Il est connecté à la veine caudale et passe en avant à travers le foie jusqu’au sinus.
Chez les embryons d’amniotes, les veines abdominales latérales sont appelées veines ombilicales ou allantoïques car elles drainent l’allantois. Les veines vitellines et ombilicales sont perdues à la naissance, seuls leurs restes persistent.
Modifications des veines:
Comme dans le cas des artères, les veines de différents vertébrés sont disposées sur le même plan fondamental. Toutes les variations montrées par eux suivent une séquence dans la série des vertébrés. Dans son développement, le système veineux des vertébrés supérieurs passe par les stades observés chez les embryons de formes inférieures.
1. Lorsque le foie se forme, la partie proximale des veines vitellines ou des veines sous-intestinales forme les veines hépatiques entre le foie et le cœur, la partie distale de la veine vitelline gauche ou de la veine sous-intestinale devient la veine porte hépatique qui forme des sinusoïdes dans le foie pour donner naissance à un système porte hépatique présent dans tous.
2. Les veines cardinales antérieures persistent sous forme de veines jugulaires internes.
3. Sauf chez les poissons, les veines cardinales communes deviennent les veines précavales qui pénètrent dans le sinus venin (amphibiens, reptiles) ou l’oreillette droite (oiseaux, mammifères). Une veine sous-clavière se forme dans chaque membre antérieur qui rejoint la veine précavière.
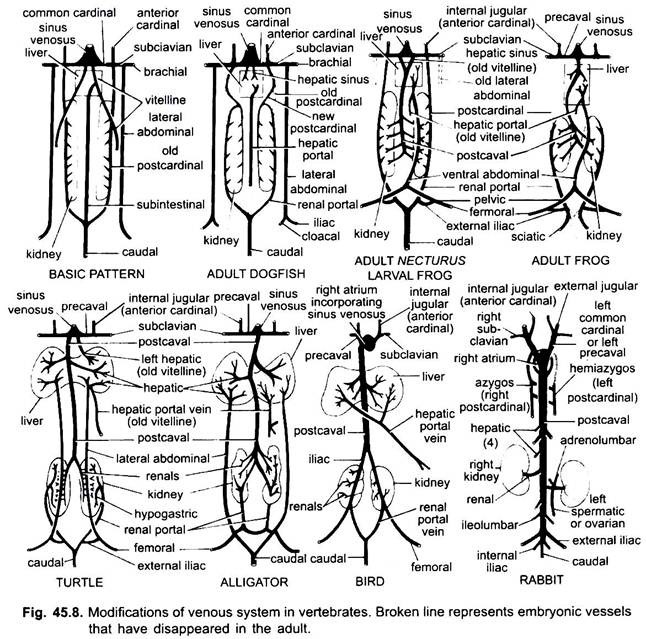
4. La veine caudale perd ses connexions avec les veines sub-intestinales et subcardinales, sa partie antérieure se divise en deux branches qui rejoignent les veines cardinales postérieures chez tous les mammifères à l’exception des mammifères.
5. Les veines cardinales postérieures persistent en tant que telles chez les poissons, mais chez d’autres, chacune se divise en deux parties, une partie antérieure qui disparaît chez les amphibiens, les reptiles et les oiseaux, mais chez les mammifères, la partie antérieure droite forme une veine azygos, la partie gauche peut former une veine hémiazygos qui perd sa connexion avec le précaval et rejoint la veine azygos par une anastomose transversale; les parties postérieures des veines cardinales postérieures qui ont rejoint la veine caudale deviennent les veines portales rénales.
La veine porte rénale de chaque côté forme des capillaires dans le rein pour former un système porte rénale qui est complet chez les poissons, les amphibiens et les reptiles, mais devient très réduit chez les oiseaux et est absent chez les mammifères car les parties postérieures des veines cardinales postérieures disparaissent chez les mammifères.
6. Les veines portales rénales des poissons ne recueillent le sang que de la queue, mais chez les amphibiens, les reptiles et les oiseaux, elles usurpent également les veines des pattes.
7. Chez certains poissons et chez tous les tétrapodes, les veines vitellines rejoignent la veine sous-cardinale pour former une veine postcavale. Chez les amphibiens et les reptiles, la veine postcavale s’étend aux extrémités postérieures des reins, chez les oiseaux, elle rejoint les veines portales rénales, réduisant ainsi le système portal rénal, chez les mammifères, la veine postcavale rejoint les veines des jambes et de la queue, de sorte que le système portal rénal est complètement éliminé.
8. Les deux veines abdominales latérales persistent en tant que telles chez les poissons, mais à partir de Dipnoï vers le haut, elles fusionnent pour former une veine abdominale antérieure qui rejoint la veine porte hépatique près du foie, la veine abdominale antérieure relie ainsi les systèmes porte rénale et porte hépatique. Une veine iliaque se forme dans chaque membre postérieur qui rejoint la veine abdominale antérieure. La veine abdominale antérieure commence à perdre de son importance chez les reptiles, chez les oiseaux elle est modifiée pour former les veines épigastriques et coccygéo-mésentériques, chez les mammifères elle disparaît, sauf chez les Tachyglossus.
9. Dans les formes respiratoires, les veines pulmonaires se développent à partir de l’oreillette gauche et pénètrent dans les poumons.
10. Les veines ombilicales (allantoïques) de l’embryon disparaissent lorsque les poumons deviennent fonctionnels en tant qu’organes respiratoires.
Tétrapodes:
Le système veineux des amphibiens ressemble beaucoup à celui des poissons-poumons, sauf que la veine abdominale pénètre dans le système portail hépatique plutôt que dans le sinus venin. Les cardinaux antérieurs persistent sous forme de veines jugulaires internes chez tous les tétrapodes adultes. Les veines jugulaires inférieures sont absentes. Les cardinaux communs deviennent la veine cave antérieure ou précavales, qui rejoignent le sinus venin chez les amphibiens et les reptiles.
Chez les oiseaux et les mammifères, les précavales pénètrent directement dans l’oreillette droite du cœur, car le sinus veneux est absent. Chez l’homme, le chat, etc., la précavale gauche est perdue de sorte que le sang du côté gauche pénètre dans la précavale droite par une branche brachio-céphalique. Chez les salamandres sans poumons, les veines pulmonaires sont absentes car le septum inter-auriculaire est incomplet.
Les veines post-cardinales rejoignent la veine caudale vers l’arrière et les précavales vers l’avant pour former les cardinaux communs chez la grenouille larvaire et le Necturus.
Chez les reptiles, il y a un plus grand développement des veines pulmonaires et des veines postcavales et une réduction de l’importance du système portail rénal qui amène le sang vers les reins depuis la partie postérieure du corps.
Chez les oiseaux, il y a deux veines précavales fonctionnelles formées par l’union de la jugulaire et de la sous-clavière de chaque côté, et une post cavale complète. Il reçoit le sang des membres par le biais des portails rénaux, qui traversent les reins, mais ne se divisent pas en capillaires et ne sont pas comparables aux portails rénaux des vertébrés inférieurs.
Chez les mammifères, il peut y avoir une ou deux veines précavales. Il existe une seule veine postcavale, dont le développement embryologique est très compliqué. Toutes les veines cavales pénètrent directement dans l’oreillette droite, car le sinus venin est absorbé dans la paroi de cette chambre du cœur dans la vie embryonnaire. Il n’existe pas de système portail rénal bien que le système portail hépatique ressemble beaucoup à celui des autres vertébrés.