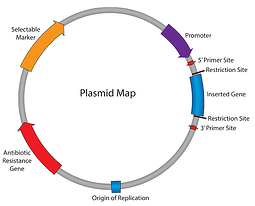
 Jusqu’à présent, dans notre série Plasmides 101, nous avons parcouru la carte des plasmides: résistance aux antibiotiques, origine de la réplication, etc. Jusqu’à ce point, nous pouvons répliquer notre plasmide et nous assurer que les cellules le maintiennent; l’étape suivante consiste à obtenir que le plasmide exprime notre gène d’intérêt. Entrez le promoteur the l’élément responsable de l’initiation de la transcription de votre insert en ARN.
Jusqu’à présent, dans notre série Plasmides 101, nous avons parcouru la carte des plasmides: résistance aux antibiotiques, origine de la réplication, etc. Jusqu’à ce point, nous pouvons répliquer notre plasmide et nous assurer que les cellules le maintiennent; l’étape suivante consiste à obtenir que le plasmide exprime notre gène d’intérêt. Entrez le promoteur the l’élément responsable de l’initiation de la transcription de votre insert en ARN.
En pratique, le terme » promoteur » décrit la combinaison du promoteur (site de liaison à l’ARN polymérase) et des opérateurs (éléments de réponse). Les promoteurs mesurent environ 100 à 1000 paires de bases et se trouvent en amont de leurs gènes cibles. La séquence de la région promotrice contrôle la liaison de l’ARN polymérase et des facteurs de transcription, les promoteurs jouent donc un rôle important dans la détermination où et quand votre gène d’intérêt sera exprimé.
La ou les ARN polymérases
sont transcrites à partir d’ADN à l’aide d’une ARN polymérase (RNAP). Chez les bactéries, cela se fait par une seule enzyme; cependant, les eucaryotes ont de multiples polymérases qui sont chacune responsables d’un sous-ensemble spécifique d’ARN. Pour acquérir cette spécificité, le RNAP eucaryote peut reconnaître et se lier à des éléments promoteurs spécifiques. Cela signifie que le promoteur présent dans votre squelette plasmidique doit être compatible avec le type d’ARN à fabriquer: si vous voulez un ARNm (pour l’expression du gène), vous devez utiliser un promoteur RNAP II, alors que les petits ARN (tels que l’ARNT) sont transcrits à partir des promoteurs RNAP III. Cet article présente les promoteurs de la transcription générale de la RNAP II et de la RNAP III; cependant, l’utilisation de LTR viraux comme promoteurs de la RNAP II est couramment utilisée dans les constructions lentivirales et rétrovirales et nous en discuterons dans un prochain article sur les parties du vecteur viral.
Spécificité du promoteur
En plus de choisir un promoteur en fonction du type de transcription de l’ARN, vous devrez également vous assurer que votre plasmide possède un promoteur adapté à votre organisme hôte. Étant donné que les mécanismes de transcription diffèrent entre les types cellulaires ou les organismes, les promoteurs doivent être également variables. Les promoteurs bactériens ne fonctionnent que dans les cellules procaryotes et généralement uniquement chez les espèces identiques ou étroitement apparentées dont ils sont dérivés. De même, les différents types de cellules eucaryotes (mammifères, levures, plantes, etc.) nécessitent des promoteurs uniques et il y a très peu de croisement. De manière générale, les promoteurs chez les bactéries sont moins diversifiés et complexes, ayant moins de parties que ceux des cellules eucaryotes. Certains promoteurs sont constitutivement actifs et actifs tout le temps tandis que d’autres sont plus soigneusement contrôlés. Les promoteurs réglementés peuvent agir uniquement dans certains tissus ou à certains moments du développement ou il peut y avoir des moyens de les allumer ou de les éteindre à volonté avec un produit chimique, de la chaleur ou de la lumière. Dans la cellule, les promoteurs eux-mêmes sont contrôlés par d’autres facteurs de régulation: des amplificateurs, des éléments limites, des isolants et des silencieux; cependant, une certaine « fuite » de la transcription peut se produire. Ce n’est normalement pas un gros problème pour les cellules, mais cela peut confondre les résultats de la recherche ou même tuer vos cellules si votre gène d’intérêt est toxique. Pour lutter contre cela, les scientifiques ont créé des promoteurs synthétiques, qui comprennent généralement une combinaison d’autres éléments promoteurs, et ont tendance à être plus étroitement réglementés.
Promoteurs communs pour les eucaryotes et les procaryotes
Nous avons inclus deux tableaux de référence ci-dessous énumérant certains des promoteurs bactériens et mammifères les plus courants. Ces listes ne sont en aucun cas exhaustives, mais devraient être un bon point de départ lorsque vous essayez de choisir votre promoteur idéal!
Promoteurs eucaryotes
| Promoteur | Principalement utilisé pour | Transcription d’ARN | Description | Expression | Considérations supplémentaires |
| CMV | Expression générale | ARNm | Promoteur d’expression mammifère fort du cytomégalovirus humain | Constitutif | Peut contenir une région activatrice. Peut être réduit au silence dans certains types de cellules. |
| EF1a | Expression générale | ARNm | Forte expression des mammifères à partir du facteur d’élongation humain 1 alpha | Constitutif | Tend à donner une expression cohérente quel que soit le type cellulaire ou la physiologie. |
| SV40 | Expression générale | ARNm | Promoteur d’expression mammifère du virus simien vacuolant 40 | Constitutif | Peut inclure un activateur. |
| PGK1 (humain ou souris) | Expression générale | ARNm | Promoteur mammalien issu du gène de la phosphoglycérate kinase. | Constitutif | Expression répandue, mais peut varier selon le type de cellule. Tend à résister à la régulation du promoteur en raison de la méthylation ou de la désacétylation. |
| Ubc | Expression générale | ARNm | Promoteur mammalien du gène de l’ubiquitine C humaine | Constitutif | Comme son nom l’indique, ce promoteur est ubiquitaire. |
| bêta-actine humaine | Expression générale | ARNm | Promoteur mammalien du gène de la bêta-actine | Constitutif | Ubiquitaire. La version poulet est couramment utilisée dans les hybrides promoteurs. |
| CAG | Expression générale | ARNm | Promoteur mammalien hybride puissant | Constitutif | Contient un activateur de CMV, un promoteur de bêta-actine de poulet et un accepteur d’épissure de bêta-globine de lapin. |
| TRE | Expression générale | ARNm | Promoteur d’élément de réponse à la tétracycline | Inductible avec la tétracyline ou ses dérivés. | Contient généralement un promoteur minimal à faible activité basale et plusieurs opérateurs de tétracycline. La transcription peut être activée ou désactivée en fonction du transactivateur tet utilisé. |
| UAS | Expression générale | ARNm | Promoteur de la Drosophile conaining des sites de liaison Gal4 | Spécifique | Nécessite la présence du gène Gal4 pour activer le promoteur. |
| Ac5 | Expression générale | ARNm | Promoteur d’insecte fort du gène de l’Actine 5c de la Drosophile | Constitutif | Couramment utilisé dans les systèmes d’expression de la Drosophile. |
| Polyèdre | Expression générale | ARNm | Promoteur fort d’insecte du baculovirus | Constitutif | Couramment utilisé dans les systèmes d’expression des cellules d’insectes. |
| CaMKIIa | Expression génique pour l’optogénétique | ARNm | Promoteur Ca2+ /calmoduline-dépendante de la protéine kinase II | Spécifique | Utilisé pour l’expression neuronale / du SNC. Modulé par le calcium et la calmoduline. |
| GAL1,10 | Expression générale | ARNm | Promoteurs adjacents de levure transcrits de manière divergente | Inductibles avec du galactose; répressibles avec du glucose | Peuvent être utilisés indépendamment ou ensemble. Réglementé par GAL4 et GAL 80. |
| TEF1 | Expression générale | ARNm | Promoteur du facteur d’élongation de transcription de levure | Constitutif | Analogue au promoteur EF1a des mammifères. |
| GDS | Expression générale | ARNm | Promoteur d’expression de levure fort à partir de la glycéraldéhyde 3-phosphage déshydrogénase | Constitutif | Très fort, également appelé TDH3 ou GAPDH. |
| ADH1 | Expression générale | ARNm | Promoteur de levure pour l’alcool déshydrogénase I | Réprimé par l’éthanol | La version pleine longueur est forte avec une expression élevée. Les promoteurs tronqués sont constitutifs avec une expression plus faible. |
| CaMV35S | Expression générale | ARNm | Promoteur végétal fort du Virus de la mosaïque du chou-fleur | Constitutif | Actif dans les dicots, moins actif dans les monocots, avec une certaine activité dans les cellules animales. |
| Ubi | Expression générale | ARNm | Promoteur végétal du gène de l’ubiquitine du maïs | Constitutif | Donne une expression élevée chez les plantes. |
| H1 | petite expression d’ARN | shRNA | Du promoteur d’ARN de la polymérase III humaine | Constitutif | Peut avoir une expression légèrement inférieure à U6. Peut avoir une meilleure expression dans les cellules neuronales. |
| U6 | petite expression d’ARN | shRNA | Du petit promoteur nucléaire U6 humain | Constitutif | U6 murin est également utilisé, mais peut être moins efficace. |
Promoteurs procaryotes
| Promoteur | Principalement utilisé pour | Description | Expression | Considérations supplémentaires |
| T7 | transcription in vitro / expression générale | Promoteur du bactériophage T7 | Constitutif, mais nécessite une ARN polymérase T7. | Lorsqu’il est utilisé pour la transcription in vitro, le promoteur pilote la transcription sensorielle OU antisens en fonction de son orientation vers votre gène. |
| T7lac | Niveaux élevés d’expression génique | Promoteur du bactériophage T7 plus opérateurs lac | Expression basale négligeable lorsqu’elle n’est pas induite. Nécessite l’ARN polymérase T7, qui est également contrôlée par l’opérateur lac. Peut être induit par IPTG. | Couramment trouvé dans les vecteurs tEP. Très étroitement réglementés par les opérateurs de bac. Bon pour moduler l’expression des gènes grâce à des concentrations d’inducteur variées. |
| Sp6 | transcription in vitro / expression générale | Promoteur du bactériophage Sp6 | Constitutif, mais nécessite une ARN polymérase SP6. | La polymérase SP6 a une processivité élevée. Lorsqu’il est utilisé pour la transcription in vitro, le promoteur pilote la transcription sensorielle OU antisens en fonction de son orientation vers votre gène. |
| araBAD | Expression générale | Promoteur de l’opéron métabolique arabinose | Inductible par la répression de l’arabinose et du catabolite réprimé en présence de glucose ou par liaison compétitive de l’anti-inducteur fucose | Plus faible. Couramment trouvé dans les vecteurs pBAD. Bon pour une régulation rapide et une faible expression basale; cependant, pas bien adapté pour moduler l’expression des gènes grâce à des concentrations d’inducteur variées. |
| trp | Niveaux élevés d’expression génique | Promoteur de E. l’opéron de tryptophane de coli | Répressible | Est désactivé avec des niveaux élevés de tryptophane cellulaire. |
| lac | Expression générale | Promoteur de l’opéron lac | Constitutif en l’absence du répresseur lac (lacI ou lacIq). Peut être induit par l’IPTG ou le lactose. | Promoteur qui fuit avec une expression quelque peu faible. La mutation lacIq augmente l’expression du répresseur 10x, resserrant ainsi la régulation du promoteur lac. Bon pour moduler l’expression des gènes grâce à des concentrations d’inducteur variées. |
| Ptac | Expression générale | Promoteur hybride de lac et de trp | Régulé comme le promoteur lac | Contient -35 région de trpB et -10 région de lac. Réglementation très stricte. Bon pour moduler l’expression des gènes grâce à des concentrations d’inducteur variées. Généralement une meilleure expression que le bac seul. |
| pL | Des niveaux élevés d’expression génique | Le promoteur du bactériophage lambda | Peut être régulé en température | Souvent associé au répresseur cI857 sensible à la température. |
| T3 | transcription in vitro / expression générale | Promoteur du bactériophage T3 | Constitutif, mais nécessite une ARN polymérase T3 | Lorsqu’il est utilisé pour la transcription in vitro, le promoteur entraîne la transcription sensorielle OU antisens en fonction de son orientation vers votre gène |
Bien que cette liste soit un excellent point de départ, les tableaux ci-dessus ne se penchent pas sur les promoteurs tissulaires ou spécifiques au développement disponibles pour les scientifiques. Les plasmides sont souvent utilisés à des fins thérapeutiques, et dans ces cas, il est important d’identifier les bons promoteurs spécifiques aux tissus, tels que décrits ici par les chercheurs du NIH.
Note : A. Max Juchheim a contribué à la rédaction de cet article.