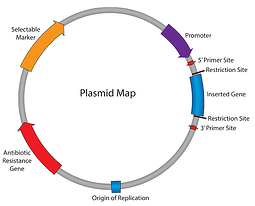
 hidtil i Vores plasmider 101-serie har vi arbejdet os gennem plasmidkortet: antibiotikaresistens, replikationens oprindelse og så videre. Indtil dette punkt kan vi replikere vores plasmid og sørge for, at celler opretholder det; det næste trin er at få plasmidet til at udtrykke vores gen af interesse. Indtast promotoren – det element, der er ansvarligt for at indlede transkriptionen af din indsats i RNA.
hidtil i Vores plasmider 101-serie har vi arbejdet os gennem plasmidkortet: antibiotikaresistens, replikationens oprindelse og så videre. Indtil dette punkt kan vi replikere vores plasmid og sørge for, at celler opretholder det; det næste trin er at få plasmidet til at udtrykke vores gen af interesse. Indtast promotoren – det element, der er ansvarligt for at indlede transkriptionen af din indsats i RNA.
i praksis beskriver udtrykket “promotor” kombinationen af promotoren (RNA-polymerasebindingsstedet) og operatørerne (responselementer). Promotorer er omkring 100 til 1000 basepar lange og findes opstrøms for deres målgener. Sekvensen af promotorregionen styrer bindingen af RNA-polymerasen og transkriptionsfaktorerne, derfor spiller promotorer en stor rolle i bestemmelsen af, hvor og hvornår dit gen af interesse vil blive udtrykt.
RNA-polymerasen(e)
RNA transkriberes fra DNA ved hjælp af en RNA-polymerase (RNAP). Eukaryoter har imidlertid flere polymeraser, som hver især er ansvarlige for en bestemt delmængde af RNA ‘ er. For at opnå denne specificitet kan den eukaryote RNAP genkende og binde til specifikke promotorelementer. Dette betyder, at promotoren, der er til stede i din plasmid-rygrad, skal være kompatibel med den type RNA, der skal laves: hvis du vil have mRNA (til genekspression), skal du bruge en RNAP II-promotor, mens små RNA ‘ er (såsom shRNA) transkriberes fra RNAP III-promotorerne. Dette indlæg indeholder promotorer til generel rnap II og RNAP III transkription; imidlertid, brug af virale ltr ‘ er som rnap II-promotorer anvendes almindeligvis i lentivirale og retrovirale konstruktioner, og vi vil diskutere disse i et fremtidigt indlæg om virale vektordele.
Promotorspecificitet
bortset fra at vælge en promotor baseret på type RNA-transkription, skal du også sørge for, at dit plasmid har en promotor, der er egnet til at arbejde i din værtsorganisme. Da transkriptionsmaskiner adskiller sig mellem celletyper eller organismer, skal promotorer være ens variable. Bakteriepromotorer virker kun i prokaryote celler og typisk kun i de samme eller nært beslægtede arter, hvorfra de blev afledt. Tilsvarende kræver de forskellige eukaryote celletyper (pattedyr, gær, planter osv.) unikke promotorer, og der er meget lidt crossover. Generelt er promotorer i bakterier mindre forskellige og komplekse og har færre dele end dem i eukaryote celler. Nogle promotorer er konstitutivt aktive og på hele tiden, mens andre er mere omhyggeligt kontrolleret. Regulerede promotorer kan kun virke i visse væv eller på bestemte tidspunkter under udvikling, eller der kan være måder at tænde eller slukke dem efter ønske med et kemikalie, varme eller lys. I cellen styres promotorer selv af stadig andre regulatoriske faktorer: forstærkere, grænseelementer, isolatorer og lyddæmpere; dog kan der forekomme en vis “lækage” af transkription. Dette er normalt ikke et stort problem for celler, men det kan forvirre forskningsresultater eller endda dræbe dine celler, hvis dit gen af interesse er giftigt. For at bekæmpe dette har forskere skabt syntetiske promotorer, som typisk inkluderer en kombination af andre promotorelementer, og har tendens til at være mere stramt reguleret.
fælles promotorer for eukaryoter og prokaryoter
vi har inkluderet to referencetabeller nedenfor, der viser nogle af de mest almindelige bakterie-og pattedyrspromotorer. Disse lister er på ingen måde udtømmende, men bør være et godt sted at starte, når du prøver at vælge din perfekte promotor!
eukaryote promotorer
| promotor | primært brugt til | RNA-transkription | beskrivelse | ekspression | yderligere overvejelser |
| CMV | generel ekspression | mRNA | stærk pattedyrekspressionspromotor fra det humane cytomegalovirus | konstitutiv | kan indeholde en forstærkerregion. Kan tavles i nogle celletyper. |
| EF1a | generel ekspression | mRNA | stærk pattedyrsekspression fra Human forlængelsesfaktor 1 alfa | konstitutiv | har tendens til at give ensartet ekspression uanset celletype eller fysiologi. |
| SV40 | generelt udtryk | mRNA | pattedyrekspressionspromotor fra simian vacuolating virus 40 | konstitutiv | kan omfatte en forstærker. |
| PGK1 (menneske eller mus) | generel ekspression | mRNA | Pattedyrspromotor fra phosphoglyceratkinasegen. | konstitutiv | udbredt ekspression, men kan variere efter celletype. Tendens til at modstå promotor ned regulering på grund af methylering eller deacetylering. |
| Ubc | generel ekspression | mRNA | Pattedyrpromotor fra det humane allestedsnærværende C-gen | konstitutiv | som navnet antyder, er denne promotor allestedsnærværende. |
| human beta actin | generel ekspression | mRNA | Pattedyrspromotor fra beta actingen | konstitutiv | allestedsnærværende. Kylling version er almindeligt anvendt i promotor hybrider. |
| CAG | generelt udtryk | mRNA | Stærk hybrid pattedyrspromotor | konstitutiv | indeholder CMV forstærker, kylling beta actin promotor og kanin beta-globin splice acceptor. |
| TRE | generel ekspression | mRNA | tetracyclin response element promotor | inducerbar med Tetracylin eller dets derivater. | indeholder typisk en minimal promotor med lav basal aktivitet og flere tetracyclinoperatører. Transkription kan tændes eller slukkes afhængigt af, hvilken tet transactivator der bruges. |
| UAS | generel ekspression | mRNA | Drosophila-promotor, der forbinder Gal4-bindingssteder | specifik | kræver tilstedeværelse af Gal4-gen for at aktivere promotor. |
| Ac5 | generel ekspression | mRNA | stærk insektpromotor fra Drosophila Actin 5C gen | konstitutiv | almindeligt anvendt i ekspressionssystemer til Drosophila. |
| Polyhedrin | generel ekspression | mRNA | stærk insektpromotor fra baculovirus | konstitutiv | almindeligt anvendt i ekspressionssystemer til insektceller. |
| CaMKIIa | genekspression for optogenetik | mRNA | Ca2+/calmodulinafhængig proteinkinase II promotor | specifik | anvendt til neuronal/CNS-ekspression. Moduleret af calcium og calmodulin. |
| GAL1, 10 | generel ekspression | mRNA | gær tilstødende, divergerende transkriberede promotorer | inducerbar med galactose; repressibel med glucose | kan anvendes uafhængigt eller sammen. Reguleret af GAL4 og GAL 80. |
| TEF1 | generelt udtryk | mRNA | gærtranskriptionsforlængelsesfaktorpromotor | konstitutiv | Analog med ef1a-promotor for pattedyr. |
| GDS | generel ekspression | mRNA | stærk gærekspressionspromotor fra glyceraldehyd 3-phosphagdehydrogenase | konstitutiv | meget stærk, også kaldet TDH3 eller GAPDH. |
| ADH1 | generelt udtryk | mRNA | Gærpromotor til alkoholdehydrogenase i | undertrykt af ethanol | fuld længde version er stærk med høj ekspression. Trunkerede promotorer er konstitutive med lavere udtryk. |
| CaMV35S | generelt udtryk | mRNA | stærk plantepromotor fra Blomkålmosaikviruset | konstitutiv | aktiv i dicots, mindre aktiv i monocots, med en vis aktivitet i dyreceller. |
| Ubi | generel ekspression | mRNA | Plantepromotor fra majs allestedsnærværende gen | konstitutiv | giver høj ekspression i planter. |
| H1 | lille RNA-ekspression | shRNA | fra den humane polymerase III RNA-promotor | konstitutiv | kan have lidt lavere ekspression end U6. Kan have bedre udtryk i neuronale celler. |
| U6 | lille RNA-ekspression | shRNA | fra den menneskelige U6 lille nukleare promotor | konstitutiv | Murine U6 bruges også, men kan være mindre effektiv. |
prokaryote promotorer
| promotor | primært brugt til | beskrivelse | udtryk | yderligere overvejelser |
| T7 | in vitro transkription/ generel ekspression | promotor fra T7 bakteriofag | konstitutiv, men kræver T7 RNA-polymerase. | når det bruges til in vitro-transkription, driver promotoren enten sense-eller antisense-transkriptet afhængigt af dets orientering til dit gen. |
| T7lac | høje niveauer af genekspression | promotor fra T7 bakteriofag plus Lac-operatører | ubetydelig basal ekspression, når den ikke induceres. Kræver T7 RNA-polymerase, som også styres af lac-operatør. Kan induceres af IPTG. | almindeligt fundet i pET-vektorer. Meget tæt reguleret af Lac-operatørerne. God til modulering af genekspression gennem forskellige inducerkoncentrationer. |
| Sp6 | in vitro transkription/ generel ekspression | promotor fra Sp6 bakteriofag | konstitutiv, men kræver SP6 RNA-polymerase. | SP6-polymerase har en høj processivitet. Når det bruges til in vitro-transkription, promotoren driver enten sense-eller antisense-transkriptet afhængigt af dets orientering til dit gen. |
| araBAD | generel ekspression | promotor af den arabinose metaboliske operon | inducerbar ved arabinose og undertrykt katabolitundertrykkelse i nærvær af glucose eller ved konkurrencedygtig binding af anti-inducer fucose | svagere. Almindeligt fundet i pBAD vektorer. God til hurtig regulering og lav basal ekspression; dog ikke velegnet til modulering af genekspression gennem forskellige inducerkoncentrationer. |
| trp | høje niveauer af genekspression | promotor fra E. coli tryptophan operon | Repressible | bliver slukket med høje niveauer af cellulær tryptophan. |
| lac | generelt udtryk | promotor fra lac operon | konstitutiv i fravær af lac-repressor (lacI eller lacik). Kan induceres af IPTG eller lactose. | utæt promotor med noget svagt udtryk. lacik mutation øger ekspression af repressoren 10 gange, således stramning regulering af lac promotor. God til modulering af genekspression gennem forskellige inducerkoncentrationer. |
| Ptac | generelt udtryk | Hybridpromotor af lac og trp | reguleret som lac-promotoren | indeholder -35 region fra trpB og -10 region fra lac. Meget stram regulering. God til modulering af genekspression gennem forskellige inducerkoncentrationer. Generelt bedre udtryk end lac alene. |
| Pl | høje niveauer af genekspression | promotor fra bakteriofag lambda | kan være temperaturregulerbar | ofte parret med den temperaturfølsomme cI857-repressor. |
| T3 | in vitro transkription / generel ekspression | promotor fra T3 bakteriofag | konstitutiv, men kræver T3 RNA-polymerase | når den bruges til in vitro transkription, driver promotoren enten sense-eller antisense-transkriptet afhængigt af dets orientering til dit gen |
selvom denne liste er et godt sted at starte, dykker tabellerne ovenfor ikke ned i det væv eller udviklingsspecifikke promotorer, der er tilgængelige for forskere. Plasmider bruges ofte til terapeutiske anvendelser, og i disse tilfælde er det vigtigt at identificere de rigtige vævsspecifikke promotorer som beskrevet af forskere ved NIH her.
Bemærk: A. maks Juchheim bidrog til skrivningen af denne artikel.