annoncer:
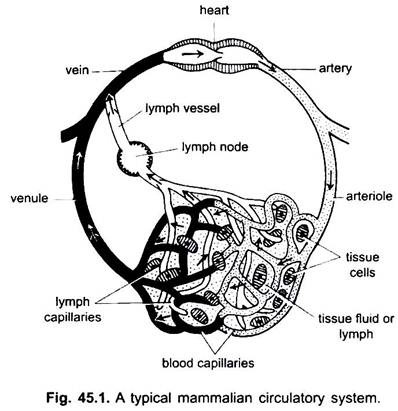
i kredsløbssystemet hos hvirveldyr er der to systemer med udførligt forgreningsrør, som forgrener sig gennem hele kroppen og bærer væsker til vævene. De er et blodvaskulært system og et lymfesystem. Når udtrykket kredsløbssystem anvendes, refererer det kun til blodvaskulærsystemet.
blodvaskulærsystemet er et lukket system hos hvirveldyr; det har et kontraktilt hjerte og kontinuerlige rør kaldet kar. Lymfesystemet er et åbent system med lymfekar. Blodkarrene, der fører blod væk fra hjertet, er arterier, der opdeles i tyndere arterioler, forgrener sig i ekstremt tynde og små kapillærer.
væggen af en kapillær er lavet af et enkelt lag af tasselerede endotelceller. Hvert blodkar, inklusive hjertet, har en foring af endotelceller eller endotel. Blod kommer ikke i kontakt direkte med vævsceller.
annoncer:
stoffer passerer fra og ind i kapillærerne gennem vævsvæsken indeholdt i vævsrum mellem celler. Udveksling af stoffer mellem kroppens blod og vævsceller finder sted gennem kapillærvæggene. Dette skyldes pinocytose i endotelcellerne i kapillærerne.
under et elektronmikroskop viser endotelcellerne mange små vesikler, som er invaginationer af plasmamembranerne. Disse vesikler bevæger sig fra den ene side af cellen og frigiver deres indhold på den modsatte side.
således giver blodet ilt, næringsstof og hormoner til cellerne for metabolisme, og cellerne giver vand, kulsyre og nitrogenholdigt affald ud i blodet for at bære dem til udskillelsesorganerne for deres hurtige eliminering.
kapillærerne danner et netværk i alle kropsvæv undtagen brusk og epitel. Fra kapillærerne passerer blodet ind i tynde venuler, der kombineres for at danne vener, der fører blod mod hjertet. Men nogle vener (portal vener, renale vener og hepatisk vene) har kapillærer, som er ligesom dem af arterier. Men alt blod passerer ikke gennem kapillærer i venuler.
annoncer:
der er også nogle gennem kanaler mellem arterioler og venuler, der ligger i nogle organer, såsom huden. Der er også arteriovenøse anastomoser mellem arterioler og venuler, der findes i cifrene. Funktionen af sådanne forbindelser er ikke klart kendt, selvom det hævdes, at de regulerer blodtryk og cirkulation.
nogle steder finder udveksling af materiale mellem væv og blod sted gennem tyndvæggede rum eller sinusoider. I visse organer kan et blodkar danne et oprullet netværk af små blodkar kaldet rete mirabile (nyrer, luftblære).
dele af kredsløbssystemet:
annoncer:
i akkordater er kredsløbssystemet af lukket type. Denne type findes også i annelider (hvirvelløse dyr). En anden type er af åben type, hvor kapillærer ikke findes. Det findes i bløddyr og leddyr. Her strømmer blodet gennem arterier ind i forskellige organer, der passerer gennem blodrum eller bihuler og derefter igen ind i kar (vener) til hjertet.
kredsløbssystemet omfatter hjerte, arterier, vener, kapillærer og blod. Hjerte er et modificeret blodkar med muskelvægge, som periodisk kontraherer for at pumpe blodet ind i de forskellige dele af kroppen gennem bestemte kar.
arterier og deres grene danner et arterielt system, der bærer blodet fra hjertet. Vener og deres bifloder udgør et venøst system, der tager blodet fra kapillærerne i arterier eller arterioler og bærer det til hjertet.
portalsystem:
i portalsystemet returneres blodet ikke direkte til hjertet, men der er et interposerende organ (lever eller nyre) i løbet af det tilbagevendende blod. Venen, der bringer blod, starter i kapillærer og ender i kapillærer, den pågældende vene fungerer både som afferente og efferente kar, de afferente kar ender i kapillærer ligesom arterier, derefter opsamles blodet i systemiske vener.
alle hvirveldyr har et hepatisk portalsystem, hvor blod passerer ind i to sæt kapillærer i leveren. Lavere hvirveldyr og embryoner fra højere hvirveldyr har et nyreportalsystem, hvor blod passerer gennem to sæt kapillærer i nyren, inden de når hjertet. Kapillærerne i hypofysen danner et hypofyseportalsystem, som er et lille, men et vigtigt system.
lymfesystem:
det findes i akkordater undtagen cyclostomer og bruskfisk. Det omfatter lymfe-og lymfekar. Lymfe er en vævsvæske, der findes blandt kroppens celler. Det er blodplasma minus røde blodlegemer og nogle proteiner.
lymfekapillærer, der danner et netværk af tynde blind-endende skibe, som samler lymfe. Lymfekar er tyndvæggede kar dannet ved forening af blodkapillærer. Disse tømmes i årer. Lymfeknuder findes på lymfekar i pattedyr. Disse danner lymfocytter af blod, der anvendes til kropsforsvar mod sygdomme.
udvikling af hjerte hos hvirveldyr:
annoncer:
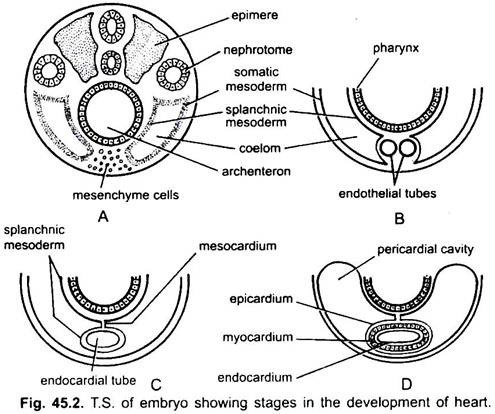
hjertet er et uparret organ, men dets oprindelse er bilateralt. I et embryo danner mesenchymet en gruppe af endokardiale celler under svælget. Disse celler bliver arrangeret til at danne et par tynde endotelrør. De to endotelrør smelter snart sammen for at danne et enkelt endokardialt rør, der ligger i længderetningen under svælget.
den splanchniske mesoderm, der ligger under endodermen, foldes i længderetningen omkring det endokardiale rør. Dette tolags rør vil danne hjertet, hvor den splanchniske mesoderm tykner for at danne et myokardium eller muskelvæg i hjertet og et ydre tyndt epikardium eller visceralt perikardium. Det endokardiale rør bliver foringen af hjertet kendt som endokardium.
folder af splanchnic mesoderm mødes ovenfor for at danne et dorsal mesokardium, der suspenderer hjertet i coelom. Snart dannes en tværgående septum bag hjertet, der deler coelom i to kamre, et forreste perikardiehulrum, der omslutter hjertet og et bageste mavehulrum.
hjertet er et lige rør, men det øges i længden og bliver S-formet, fordi dets ender er faste. Udseende af ventiler, indsnævring, skillevægge i hjertet og differentielle fortykkelser af dets vægge danner tre eller fire kamre i hjertet.
1. Single-Chambered Heart:
i amphioksus (primitivt akkordat) findes et ægte hjerte ikke. En del af ventral aorta under svælget er muskuløs og kontraktil og fungerer som hjerte.
2. To-Kammer Hjerte:
i cyclostomer er der fire kamre arrangeret i en lineær rækkefølge – en tyndvægget sinus venosus, et let muskulært atrium (auricle), en muskuløs ventrikel og en muskuløs Conus arteriosus eller bulbus cordis. Det ligger i kropshulrummet, hvor andre viscerale organer også er til stede.
ud af fire kamre svarer kun atrium og ventrikel til de fire kamre (parrede atria og parrede ventrikler) hos de højere hvirveldyr. I hjertets udvikling har mange ændringer fundet sted.
Elasmobranchs:
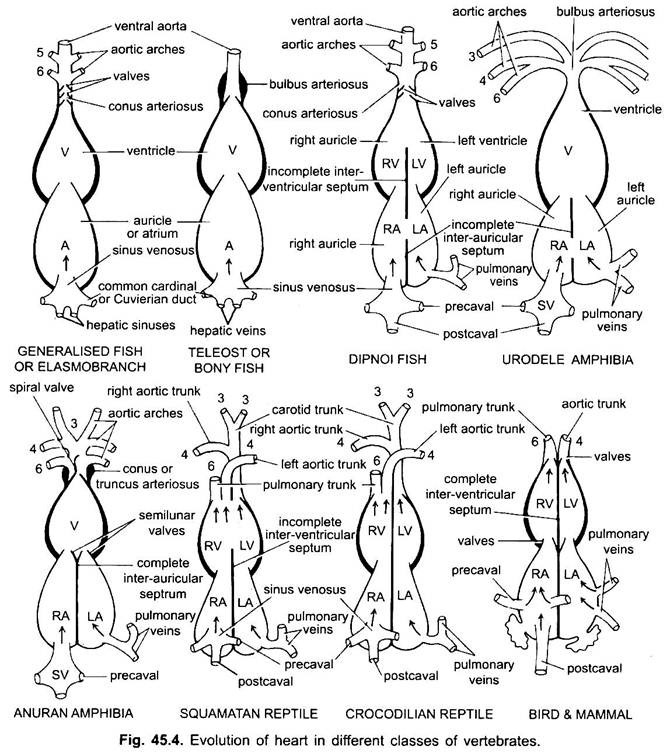
undtagen Dipnoi, kredsløbssystemet i fisk fra cyclostomer til teleosts, går kun uilt blod til hjertet, derfra pumpes det til gællerne, luftes og fordeles derefter til kroppen. Hjertet af bruskhundfisk er muskuløst og dorsoventralt bøjet S-formet rør med fire rum i en lineær serie.
de er sinus venosus og atrium til modtagelse af venøst blod og en ventrikel og conus arteriosus til pumpning af dette blod. Hjertet er et forgrenet venøst hjerte. Sinus venosus og Conus arteriosus er tilbehørskamre. Atrium og ventrikel er sande kamre, således er det et 2-kammer hjerte.
sinus venosus åbner anteriort ind i atrium gennem sinu-atrial blænde beskyttet af et par ventiler. Atrium ligger dorsal til ventrikel og åbner ventralt ind i ventrikel gennem en atrio-ventrikulær blænde beskyttet af et par ventiler. Den tykvæggede, muskulære ventrikel åbner ind i en smal conus arteriosus indeholdende ventiler i to serier.
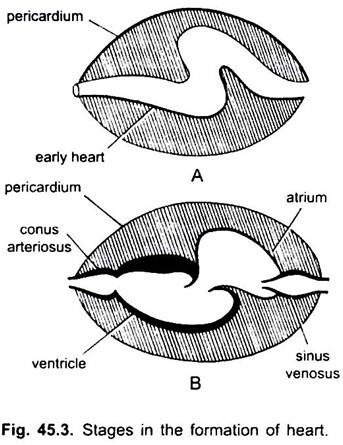
hjertet er indesluttet i perikardial hulrum adskilt fra kropshulrum ved en tværgående septum. Conus gennemborer perikardiet og bliver kontinuerligt med den ventrale aorta. Perikardial hulrum kommunikerer med kropshulrummet gennem to perforeringer i den tværgående septum.
Teleosts:
deres hjerte ligner clasmobranchs. I teleosts er conus reduceret og har et enkelt par ventiler. Den proksimale del af ventral aorta tæt på conus bliver stærkt forstørret og tykvægget, kaldet bulbus arteriosus. Det er elastisk og udvides på tidspunktet for ventrikulær sammentrækning. Hjertet er således 2-kammeret med en enkelt blodcirkulation.
3. Tre-kammer hjerte:
Dipnoi:
i diphoans deler en septum atriumet i et højre og venstre kammer. Dette er korreleret med brugen af svømmeblæren som et åndedrætsorgan og repræsenterer det første skridt mod udviklingen af det dobbelte kredsløbssystem, hvorved både iltet og uilt blod kommer ind i hjertet og holdes adskilt.
blod fra lungfiskens højre Aurikel passerer ind i højre ventrikel og pumpes derefter ind i den primitive lungelignende gasblære af lungearterier, der forgrener sig fra det sjette par aortabuer. Det iltede blod vender tilbage til venstre atrium ved hjælp af lungeårer som amfibier.
Amphibia:
i amfibier skifter dorsal atrium anterior til ventrikel. Sinus venosus åbner ind i højre atrium dorsalt og ikke bagved. Atriet er fuldstændigt opdelt i højre og venstre kamre og har ingen foramen ovale i den inter-aurikulære septum, som forbliver åben i dipnoans.
dybe lommer udvikler sig i det ventrikulære hulrum. Conus arteriosus opdeles i systemiske og pulmonale kar ved hjælp af en spiralventil. I lunge mindre salamandere er den interatriale septum ufuldstændig, og lungevene er fraværende.
Reptilia:
i krybdyr er hjertet yderligere avanceret. Atriet er altid fuldstændigt adskilt i et højre og venstre kammer, og i mange former er sinus venosus inkorporeret i væggen i det højre atrium. Ventriklen er også delvist opdelt af en septum i de fleste krybdyr, og i alligatorer og krokodiller er helt to-kammer.
dette betyder, at iltet blod, der kommer fra lungerne til venstre side af hjertet, i det væsentlige adskilles fra det ikke-iltede blod fra kroppen til højre side. I krokodiller er de to blodtyper således fuldstændigt adskilt og næsten komplette i andre krybdyr, men en vis blanding forekommer i andre dele af kredsløbssystemet.
den embryonale conus arteriosus opdeles i tre i stedet for to kar:
(i) Lungebue, der bærer blod til lungerne fra højre side af ventriklen.
(ii) højre systemisk aorta, der bærer blod fra venstre side af ventriklen til kroppen ved hjælp af højre fjerde aortabue.
(iii) venstre systemisk kommer fra højre ventrikel til venstre fjerde aortabue.
ved kontaktpunktet med den systemiske aorta fra venstre ventrikel, selv hos krokodiller, er der en åbning mellem de to, kaldet foramen af Panise, hvor der kan være en vis blanding af de to typer blod. Således repræsenterer reptilhjerte overgangshjerte mod amfibiehjerte-2 komplette aurikler og 2 ufuldstændige ventrikler med en lille blanding af blod i højre og venstre systemisk.
4. Fire-kammer hjerte:
Aves og Mammalia:
hos fugle er ventriklen helt opdelt i to, så hjertet er fire kammer (2 aurikler og 2 ventrikler). Der er fuldstændig adskillelse af venøst og arterielt blod. Den systemiske aorta forlader venstre ventrikel og fører blod til hoved og krop. Mens lungearterien forlader højre ventrikel og fører blod til lungerne for iltning.
der er således dobbelt cirkulation, hvor der ikke er nogen blanding af blod på noget sted. Sinus venosus er fuldstændigt inkorporeret i højre auricle, som modtager to precavals og en postcaval. Den venstre Aurikel modtager iltet blod gennem lungevene, conus arteriosus er fraværende, pulmonal aorta opstår fra højre ventrikel, og enkelt systemisk aorta opstår fra venstre ventrikel, og begge har ventiler ved deres baser.
modifikationer af Aortabuer hos hvirveldyr:
embryonale arterier:
når hjertet dannes i et hvirveldyrsembryo, vises et blodkar kaldet ventral aorta midt-ventralt under svælget, som snart bliver forbundet med conus arteriosus. Den ventrale aorta, der opstår fra hjertet, løber fremad under svælget og deler sig anteriorly i et par ydre halspulsårer i hovedet.
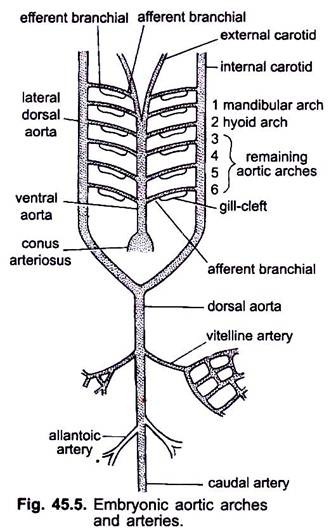
den ventrale aorta afgiver 6 par laterale aortabuer ved ækvivalens, der løber gennem de viscerale buer. Hver aortabue har en ventral afferent grenarterie, der bærer venøst blod til gællen og en dorsal efferent grenarterie, der tager iltet blod fra gællen. De efferente grenarterier på begge sider forbinder dorsalt med den laterale dorsale aorta eller Radik aorta, der kommer ind i hovedet som indre halspulsårer.
den første aortabue er en mandibulær aortabue, den anden er en hyoid aortabue, de resterende kaldes tredje, fjerde, femte og sjette aortabuer. Den laterale dossale aorta smelter bag svælget for at danne en dorsal aorta, der fortsættes midt-dorsalt ind i halen som en kaudal arterie.
fra dorsal aorta opstår parrede og uparrede arterier, som leverer forskellige organer i kroppen. I et embryo med en blommesæk opstår et par vitellinarterier fra dorsal aorta og leverer blommesækken. I embryoner af amnioter opstår et par navlestrengs-eller allantoiske arterier fra dorsal aorta, der leverer blod til allantois.
hos en voksen smelter vitellinarterierne sammen for at danne den vigtigste mesenteriske arterie, hovedparten af de allantoiske arterier går tabt, men deres rester danner hypogastriske eller indre iliacarterier.
Aortabuer hos hvirveldyr:
hos forskellige voksne hvirveldyr ser det arterielle system ud til at være anderledes, men de er bygget på den samme grundlæggende grundlæggende plan. Forskellen skyldes stigende kompleksitet i hjertet på grund af en ændring i respiration fra gæller til lunger. Der er en progressiv reduktion i antallet af aortabuer i hvirveldyrserien.
Cyclostomata:
i Petromyson er der 7 par aortabuer. I andre cyclostomer varierer disse fra 6 par i Eptatretus og 15 Par i Eptatretus.
Fiskene:
selvom seks anses for at have været det grundlæggende antal aortabuer for fisk. Dette tal reduceres til fem selv i hajer og stråler med tabet af det første par, den mandibulære aortabue eller den er repræsenteret af en efferent pseudobranchial arterie. I de fleste benede fisk forsvinder både mandibular (i) og hyoid (ii) aortabuer eller er meget reduceret.
hos Polypterus og dipnoi (lungefisk) er gæller ikke veludviklede. Så lungearterien stammer fra den efferente del af 6.bue på begge sider, der leverer blod til luftblæren eller lungen. I elasmobranchs og Dipnoi har hver bue en afferente og to efferente grenarterier (dannet ved opdeling) i hver gill. Hos benede fisk har hver gælle en afferent og en efferent arterie.
i tetrapoda bryder aortabuer ikke op i afferente og efferente dele, fordi ægte indre gæller er fraværende. I alle tetrapoda forsvinder den første og anden bue.
Amphibia:
her viser aortabuerne modifikation på grund af tab af gæller og udseende af lungerne. I urodeler er der eksterne gæller til stede som åndedrætsorganer ud over lungerne. III, IV, V og VI aortabuer er til stede, selvom det femte par er meget reduceret i Sirene, Amphiuma og necturus. Aortabuerne brydes ikke i de ydre gæller i afferente og efferente dele, fordi grene, der stammer fra IV, V og VI aortabuer, danner kapillærer i de ydre gæller.
den laterale dorsale aortae mellem III og IV aortabuer vedvarer som en vaskulær forbindelse, ductus caroticus. VI aortabuen danner den pulmo-kutane bue eller arterie på begge sider, der tager blod til lungen og huden. Det bevarer også en forbindelse med den laterale dorsale aorta kendt som en ductus arteriosus (kanal af Botalli).
i larven af anuran (frøen tadpole) er arrangement af aortabuer som en voksen urodele på grund af tilstedeværelsen af gæller. Ved metamorfose, med tab af gæller, forsvinder i, II og V aortabuer fuldstændigt, kun iiird, IVth og Vlth aortabuer er til stede. Den laterale dorsale aorta mellem den tredje og fjerde aortabue (ductus caroticus) forsvinder også. Således bliver den tredje aortabue sammen med en del af den ventrale aorta carotisbuen, der bærer iltet blod til hovedområdet.
den fjerde aortabue langs dens laterale dorsale aorta danner den systemiske bue. Den sjette aortabue bliver den pulmocutane bue, der leverer venøst blod til lunger og hud. Ductus arteriosus forsvinder under metamorfose. Således har voksne anuraner kun III, IV og VI aortabuer. Disse bevares også af amnioter.
Reptilia:
i reptiler erstattes gællerne fuldt ud af lunger. Kun III, IV og VI aortabuer er til stede. Med den delvise adskillelse af ventriklen i to dele opdeles den distale del af conus arteriosus og hele den ventrale aorta i tre kar, dvs.to aorta eller systemiske og en lunge. Højre systemisk bue (IV) stammer fra venstre ventrikel, der bærer iltet blod til carotidbuen (III). Den venstre systemiske (IV) og pulmonale aortae (VI) stammer fra højre ventrikel. Den venstre systemiske bærer afilt eller blandet blod til kroppen gennem dorsal aorta.
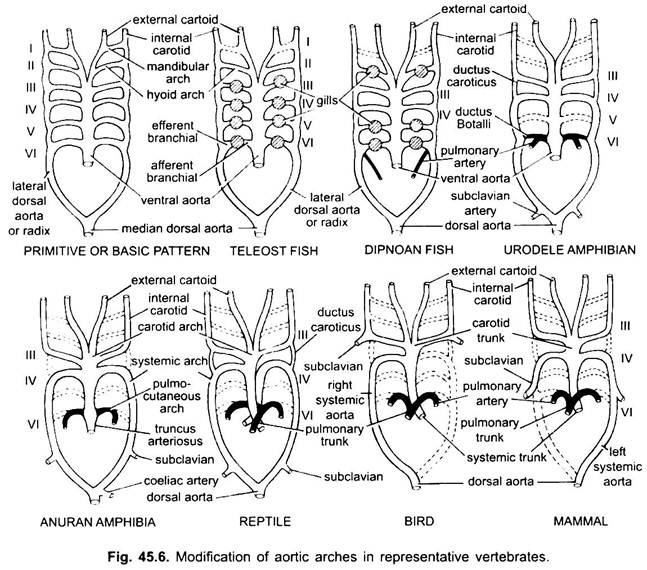
mens lungearterien tager afilt blod til lungerne. Ductus caroticus forsvinder, men det fortsætter i slanger og nogle firben (Uromastiks). Ductus arteriosus forsvinder i de fleste krybdyr, selvom den vedvarer i en reduceret form i Sphenodon og nogle skildpadder. På grund af blanding af blod er krybdyr koldblodede dyr som fisk og amfibier.
Aves:
hos fugle er III, IV og VI aortabuer til stede. De følger det generelle mønster af krybdyr med nogle forskelle. Med den komplette opdeling af ventriklen i to dele er conus arteriosus og ventral aorta opdelt for at danne to kar, systemisk aorta, der stammer fra venstre ventrikel og en pulmonal aorta fra højre ventrikel.
tredje aortabue med rester af lateral og ventral aortae danner carotiderne, der opstår fra systemisk aorta. Fjerde aortabue danner kun den systemiske aorta på højre side. Det forener sig med den laterale aorta på sin egen side og danner dorsal aorta. En del af den fjerde aortabue på venstre side danner den venstre subklaviske arterie, resten sammen med dens laterale dorsale aorta forsvinder. Den sjette aortabue danner pulmonal aorta. Ductus caroticus og ductus arteriosus forsvinder.
Mammalia:
hos pattedyr vedvarer også III, IV og VI aoriske buer. Ventriklen er opdelt fuldstændigt i to dele.
conus arteriosus og ventral aorta splittede sig for at danne to kar:
(i) en systemisk aorta, der stammer fra venstre ventrikel, og
(ii) en pulmonal aorta fra højre ventrikel. Tredje aortabue med rester af lateral og ventral aortae danner carotidbuen.
fjerde aortabue danner kun den systemiske aorta på venstre side, mens den på højre side danner den proksimale del en innomineret og højre subclavian arterie, resten sammen med dens laterale dorsale aorta forsvinder. Sjette aortabue danner pulmonal aorta. Ductus arteriosus degenererer, men den vedvarer i nogle indtil udklækning eller fødsel i reduceret form på venstre side som et tyndt ligamentum arteriosum.
Venøst System:
Embryonale Vener:
i alle hvirveldyrsembryoer er venesystemet enkelt og lignende. Venerne er for det meste parret og symmetrisk arrangeret. I embryoner uden en æggeblomme sac dannes en sub – intestinal vene i splanchnic mesoderm under tarmen.
den løber rundt om anus og fortsættes bagud som:
(i) en kaudal vene i halen. I alle embryoner, der har en æggeblomme (uanset om de indeholder æggeblomme eller ej), opstår et par vitellinårer fra æggeblommesækken og slutter sig til den bageste del af det udviklende hjerte, der bliver sinus venosus. Faktisk er fusionen af disse vitellinårer ansvarlig for hjertedannelsen i benede fisk, krybdyr og fugle. Hver vitellin vene i sin bageste ende forbinder den subintestinale vene dannet på samme måde som i embryonerne uden æggeblomme.
(ii) et par subkardinale vener opstår mellem nyrerne og slutter sig til kaudalvenen.
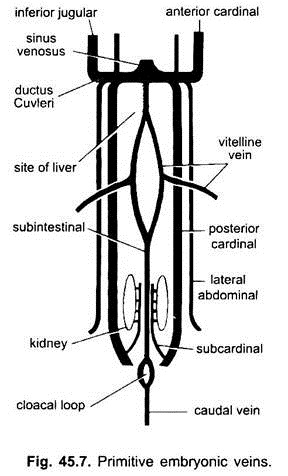
(iii) parrede forreste og bageste kardinalårer dannes, som bærer blodet fra henholdsvis hovedet og de bageste dele af kroppen. De forreste og bageste kardinalvener på hver side forenes for at danne en ductus Cuvieri eller fælles kardinalven, der passerer indad gennem den tværgående septum for at komme ind i sinus venosus.
hos fisk og salamandere (urodeles) kommer en ringere jugular ven fra den ventrale side af hovedet for at slutte sig til den fælles kardinalven. Det har ingen homolog hos andre hvirveldyr. I en amniotes kommer et par laterale eller ventrale abdominale vener fra kropsvæggen for at komme ind i de fælles kardinalårer.
Fiskene:
almindelig kardinalven (kanal af Cuvier) kommer ind i sinus venosus fra hver side og dannes ved fusion af forreste og bageste kardinaler. Blod fra hovedet opsamles af de forreste kardinaler, og blod fra nyrerne og gonaderne opsamles af de bageste kardinaler.
også ind i kanalerne i Cuvier er de parrede laterale abdominale vener, der modtager blod fra kropsvæggen og parrede vedhæng. Det renale portalsystem består af den kaudale vene og to renale portalvener placeret lateralt til nyrerne, som kapillariserede i nyrerne.
hepatisk portalsystem bærer blod fra maven og tarmen og returnerer det til leveren, hvor det efter at have passeret en række sinusoider kommer ind i sinus venosus ved hjælp af parrede hapatiske vener.
i teleosts mangler de laterale abdominale vener, således at blod fra subclavians, dræning af pectoral appendages, kommer direkte ind i sinus venosus, og blodet fra iliac vener, der dræner bækkenvedhængene, passerer ind i postkardinalerne.
i dipnoans er en enkelt ventral abdominal vene til stede, formodentlig afledt af en fusion af laterale abdominaler. Denne vene modtager blod fra ilacs ved hjælp af parrede bækkenårer og kommer ind i den højre kanal af Cuvier. Fra det højre postkardinale system, en ny vene, ser postclavalen desuden ud, som er af stor betydning hos højere hvirveldyr. Det er forbundet med den kaudale vene og passerer fremad gennem leveren til sinus.
i embryoner af amnioter er de laterale abdominale vener kendt som navlestreng eller allantoiske vener, fordi de dræner allantois. Vitellinen og navlestrengen går tabt ved fødslen, kun deres rester vedvarer.
modifikationer af vener:
som i tilfælde af arterier er vener af forskellige hvirveldyr arrangeret på samme grundlæggende plan. Eventuelle variationer vist af dem følger en sekvens i hvirveldyrserien. I sin udvikling passerer det venøse system af højere hvirveldyr gennem de stadier, der ses i embryoner af lavere former.
1. Når leveren dannes den proksimale del af vitellinvenerne eller subintestinale vener danner levervenerne mellem leveren og hjertet, bliver den distale del af venstre vitellinven eller subintestinalvenen den hepatiske portalven, der danner sinusoider i leveren for at give anledning til et hepatisk portalsystem, der er til stede i alle.
2. Forreste kardinal vener vedvarer som indre jugular vener.
3. Undtagen hos fisk bliver de almindelige kardinalårer de prækavale vener, der kommer ind i sinus venosus (amfibier, krybdyr) eller den højre Aurikel (fugle, pattedyr). En subclavian vene dannes i hver forben, der forbinder den prækavale vene.
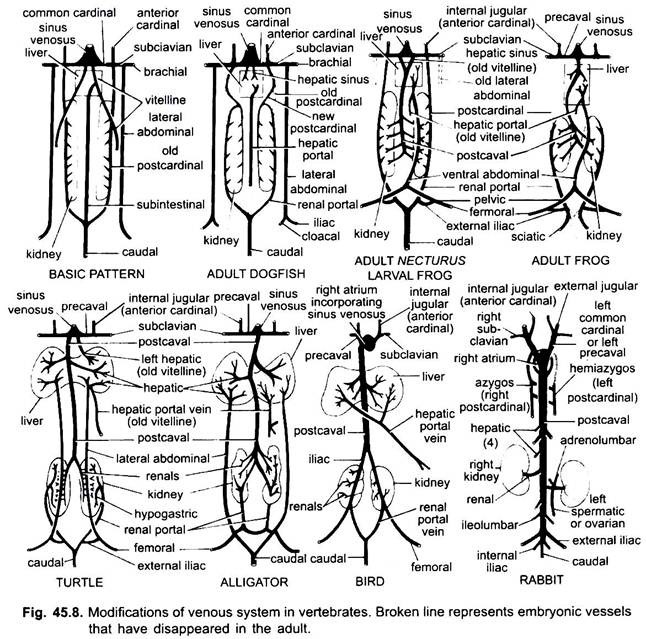
4. Den kaudale vene mister sine forbindelser med de subintestinale og subkardinale vener, dens forreste del opdeles i to grene, der forbinder de bageste kardinalårer i alle undtagen pattedyr.
5. De bagerste kardinalvener består som sådan hos fisk, men hos andre opdeles hver i to dele, en forreste del, der forsvinder hos padder, krybdyr og fugle, men hos pattedyr danner den højre forreste del en aygosven, den venstre kan danne en hemiasygosven, der mister sin forbindelse med prækavalen og slutter sig til asigosvenen ved en tværgående anastomose; de bageste dele af de bageste kardinalvener, der har sluttet sig til kaudalvenen, bliver de renale portalvener.
nyreportalvenen på hver side danner kapillærer i nyren til dannelse af et nyreportalsystem, der er komplet hos fisk, padder og krybdyr, men bliver meget reduceret hos fugle og er fraværende hos pattedyr, fordi de bageste dele af bageste kardinalårer forsvinder hos pattedyr.
6. Nyreportalvenerne af fisk samler kun blod fra halen, men i amfibier, krybdyr og fugle bruger de også venerne fra benene.
7. Hos nogle fisk og alle tetrapoda slutter vitelline-venerne sig til den subkardinale vene for at danne en postkaval vene. Hos padder og krybdyr strækker postkavalvenen sig til de bageste ender af nyrerne, hos fugle slutter den sig til nyreportalvenerne, hvilket reducerer nyreportalsystemet, hos pattedyr forbinder postkavalvenen venerne fra benene og halen, så nyreportalsystemet elimineres fuldstændigt.
8. De to laterale abdominale vener vedvarer som sådan hos fisk, men fra Dipnoi opad smelter de sammen for at danne en anterior abdominal vene, der forbinder leverportalvenen nær leveren, den forreste abdominalven forbinder således nyreportalen og leverportalsystemerne. En iliac vene dannes i hver bagben, der forbinder den forreste abdominale vene. Den forreste abdominale vene begynder at miste sin betydning i krybdyr, hos fugle er den modificeret til at danne de epigastriske og coccygeo-mesenteriske vener, hos pattedyr forsvinder den, undtagen i Tachyglossus.
9. I luftpustende former vokser lungevene ud fra venstre auricle og kommer ind i lungerne.
10. Embryoets umbilical (allantoic) vener forsvinder, når lungerne bliver funktionelle som åndedrætsorganer.
Tetrapoda:
det venøse system af amfibier ligner meget lungfiskene, bortset fra at abdominalvenen kommer ind i leverportalsystemet snarere end sinus venosus. Forreste kardinaler vedvarer som indre jugular vener i alle voksne tetrapoder. Ringere jugular vener er fraværende. Almindelige kardinaler bliver de forreste vena cavae eller precavals, som forbinder sinus venosus i amfibier og krybdyr.
hos fugle og pattedyr kommer prækavaler direkte ind i højre Aurikel i hjertet, fordi sinus venosus er fraværende. I mand, kat osv., venstre precaval går tabt, så blodet fra venstre side kommer ind i højre precaval gennem en brachio-cephalisk gren. I lungeløse salamandere er lungevene fraværende, da inter-atrial septum er ufuldstændig.
post-kardinalvenerne slutter sig til kaudalvenen bagved og precavals anteriorly for at danne de fælles kardinaler i larvefrøen og Necturus.
hos krybdyr er der større udvikling af lungeårer og postkaval vene og reduktion i vigtigheden af nyreportalsystem, der bringer blod til nyrerne fra den bageste del af kroppen.
hos fugle er der to funktionelle prækavale vener dannet ved forening af jugular og subclavian på hver side og en komplet postkaval. Det modtager blod fra lemmerne ved hjælp af nyreportalerne, der passerer gennem nyrerne, men bryder ikke op i kapillærer og kan ikke sammenlignes med nyreportalerne hos lavere hvirveldyr.
hos pattedyr kan der være en eller to prækavale vener. Der er en enkelt postkaval vene, hvis embryologiske udvikling er meget kompliceret. Alle kavale vener kommer direkte ind i højre Aurikel, da sinus venosus absorberes i væggen i dette hjertekammer i embryonalt liv. Der er ikke noget nyreportalsystem, selvom leverportalsystemet meget ligner det hos andre hvirveldyr.