- introduktion
- resultater
- Dbh-Cre-driveren markerer og er begrænset til th-udtrykkende noradrenerge regioner i hjernestammen, der hæmmes af CNO-Administration
- CNO-hM4D-medieret forstyrrelse af noradrenerge neuroner hos voksne mus
- CNO-hM4D forstyrrelse af Dbh-Cre neuroner resulterer i et hyperkapnisk underskud
- Habituerede Søskendekontrol injiceret med 10 mg / kg CNO viste et hyperkapnisk Ventilationsunderskud
- Habituated og Unhabituated vildtype Dyr injiceret med saltvand viste ingen ændringer Presalin og Postsaline
- koncentrationsforhold mellem Closapin og CNO er højere i hjernen end i Serum
- Diskussion
- materialer og metoder
- etisk godkendelse
- avl, genetisk baggrund og vedligeholdelse af mus
- histologi
- elektrofysiologi
- plethysmografi
- væskekromatografi-massespektrometri
- dataindsamling og analyse
- plethysmografi
- statistik
- plethysmografi
- væskekromatografi-massespektrometri
- Etikerklæring
- Forfatterbidrag
- finansiering
- interessekonflikt Erklæring
- anerkendelser
introduktion
de neurale netværk, der ligger til grund for den respiratoriske kemosensoriske refleks, er et primært mål for forståelse af etiologien af flere adfærdsmæssige og fysiologiske lidelser. Forstyrrede kemosensoriske reflekser antages at spille en rolle i både medfødte og voksne lidelser, herunder pludseligt Spædbarnsdødssyndrom (SIDS), medfødt centralt Hypoventilationssyndrom (CCHS), centrale søvnapnøer, unormal vejrtrækning i Rett syndrom og fedme hypoventilationssyndrom (Obonai et al., 1998; Jens et al., 2003; Amiel et al., 2009; Ramanantsoa et al., 2011; et al., 2013). Under hypotesen om falsk kvælningsalarm menes upassende kemosensorisk aktivering eller overfølsomhed at spille en rolle i undergrupper af patienter, der lider af panikforstyrrelse (Klein, 1993). Kemosensorisk dysfunktion kan også spille en rolle i neurodegenerative sygdomme gennem søvnforstyrret vejrtrækning, der er forbundet med accelereret progression (Hakim et al., 2012; Verbraecken og McNicholas, 2013; Bahia og Pereira, 2015; Snyder et al., 2017). Således vil en bedre forståelse af hjernestammen kemosensoriske netværk give vigtige spor til en række adfærdsmæssige og fysiologiske patologier.
farmakogenetiske designerreceptorer udelukkende aktiveret af designer drugs (DREADDs) (Armbruster et al., 2007) har været anvendt i en række undersøgelser for at kortlægge neurale populationer i respiratorisk kemorefleks (åndedrætsrespons på forhøjede CO2-niveauer i blodet). DREADD-teknologi i kombination med tværsnitsgenetisk implementering er blevet brugt til at dæmpe meget målrettede neuronale populationer, mens vi observerer åndedrætsfunktion i bevidste og uhæmmede mus af vores laboratorium og andre og undgår mange forvirringer fra tidligere kredsløbskortlægningsmetoder (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Disse og de fleste andre undersøgelser omfattede næsten altid CNO kun ikke-DREADD, der udtrykte søskendekontrol, der ikke viste kemosensoriske eller andre respiratoriske virkninger, idet de argumenterede for, at CNO ikke havde nogen off-target-virkninger på vejrtrækning i bevidste og uhæmmede mus på trods af den høje dosis, der blev brugt, 10 mg/kg. Ikke desto mindre blev det fundet i flere andre undersøgelser, at CNO og dets rygmetabolismeprodukter kunne have off-target virkninger på adfærd og bevægelse i en række analyser, men respiratorisk output blev ikke behandlet (Guettier et al., 2009; Joober, 2010; MacLaren et al., 2016; Gomes et al., 2017; Ilg et al., 2018; Mahler og Aston-Jones, 2018; Manvich et al., 2018; Padovan og Knackstedt, 2018). Det blev også vist, at CNO og dets metabolitter ikke er ækvivalent fordelt mellem kredsløbssystemet og hjernen., 2017).
en anden, uadresseret bekymring stammer fra den stress, der er induceret af det eksperimentelle paradigme, herunder at holde dyr i et barometrisk plethysmografikammer i hele kroppen, håndtering, intra-peritoneal injektion og rektal temperaturmålinger. I tidligere dreadd-åndedrætsundersøgelser blev na-kurvemus introduceret i et lille kammer (140-400 mls) og fik lov til at akklimatisere 20-40 minutter før dataindsamling (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Det er imidlertid ikke klart, om denne tid er tilstrækkelig til at minimere stress-inducerede respiratoriske ændringer, der kan fungere som en interagerende faktor med virkningerne af closapin.
i vores undersøgelser for at undersøge det noradrenerge (NA) systems rolle i respiratorisk kontrol forsøgte vi at anvende den veletablerede RC::P_HM4D DREADD allel (Ray et al., 2011) for at teste na-neurons rolle i baseline og hyperkapnisk respiration hos uhæmmede voksne dyr ved hjælp af helkropsplethysmografi. Fordi noradrenalin og NA neuroner også er kendt for at spille en central rolle i stressresponser (Valentino og Van Bockstaele, 2008; Chen et al., 2018), gennemførte vi en række undersøgelser for at sammenligne omfattende habituation (flere eksponeringer for det eksperimentelle paradigme, hvorved dyret lærer at oplevelsen er ikke-truende eller overlevelig) og høj vs. lav (1 mg/kg) CNO doser med tidligere offentliggjorte DREADD protokoller (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Her viser vi for første gang, at høje systemiske doser af CNO er i stand til at fremkalde off-target-effekter på en autonom åndedrætsfunktion hos bevidste mus. Vi afslører også, at CNO ‘ s off-target-effekt på kemosensorisk åndedrætsudgang effektivt afmaskeres ved omfattende tilvænning og derfor ikke ville være tydelig i tidligere CNO-kontrolundersøgelser, der ikke habituerede dyr før åndedrætsmåling, idet der kun anvendes en kort akklimatiseringsperiode inden dataindsamling. Tilsammen antyder disse resultater, at tidligere kortlagte neuronale populationer indirekte kan påvirke respiratorisk kontrol gennem potentielle roller i regulering af stressresponser. Disse data stemmer især overens med nylige rapporter, der tyder på, at CNO ikke er biologisk inert ved høje doser via metabolisk omdannelse til Maclaren et al., 2016; Gomes et al., 2017), og at adfærdseffekter uden for målet kan manifestere sig ikke kun fra forstyrrelse af adfærdskredsløb, men også fra forstyrrelser til underliggende autonome kredsløb og homeostase.
resultater
Dbh-Cre-driveren markerer og er begrænset til th-udtrykkende noradrenerge regioner i hjernestammen, der hæmmes af CNO-Administration
for at undersøge udtrykket og specificiteten af DBH-Cre-linjen brugte vi en enkelt rekombinase avlsordning (figur 1a), hvor vi krydsede føreren til AI9-linjen (Madisen et al., 2010), som udtrykker en flokset tdTomato. Tdtomat i hjernestammen var begrænset til th-udtrykkende regioner, herunder locus coeruleus, A5, A1, A2, A7, SubCV og SubCD kerner som forventet (figur 1b). For at bekræfte, at NA-neuroner, der udtrykker hM4D-receptoren, reagerede på CNO, udførte vi optagelser på locus coeruleus (LC) neuroner, hvor vi observerede en inhibering af neuronfyring ved CNO-badapplikation (n = 3, figur 1C).
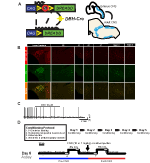
Figur 1. Avl skematiske og respiratoriske protokoller. RC::P_hM4D-allel kombineres med en dopamin-beta-hydroksylase (DBH)-Cre-linje for at opnå cellespecifik ekspression af hM4D-receptoren i noradrenerge (NA) neuroner (a). Fluorescerende ekspression af tdtomat i DBH-Cre; AI9 koronal hjerne sektioner Co-udtrykker med tyrosin-hydroksylase (TH) i hjernestammen NA kerner (B). I Skive optagelse af NA locus coeruleus før og efter CNO ansøgning, demonstrerer DREADD receptor funktionalitet i målrettede neuroner (C). Respiratorisk protokol. Habituerede mus udsættes for en 5-dages habitueret protokol bestående af håndtering, rektal sondetemperaturmåling, saltinjektion og eksponering for plethysmografi-kammeret. På den sjette dag, både habituated og unhabituated mus under et hyperkapnisk assay, hvor dyret placeres i åndedrætskammeret og får lov til at akklimatisere under baseline rumluftforhold. Dyret udsættes derefter for 20 min 5% CO2 efterfulgt af 20 min rumluft. Dyret injiceres derefter intraperitonealt med CNO eller saltvand efterfulgt af yderligere 20 minutter rumluft efter injektion, 20 minutter på 5% CO2 og 20 minutter rumluft. Åbne kasser afgrænser dataindsamlingstider (D).
CNO-hM4D-medieret forstyrrelse af noradrenerge neuroner hos voksne mus
for at undersøge rollen af NA-neuroner under baseline og hyperkapnisk respiration anvendte vi RC:P_hM4D-hæmmende DREADD-system krydset med DBH-Cre-driveren. Brug af helkropsplethysmografi (Ray et al., 2011), målte vi ventilationsresponserne hos uhæmmede voksne dyr under rumluft (21% O2/79% N2) og hypercapnic (5% CO2/21% O2/74% N2) betingelser før og efter CNO-administration (figur 1D). For at imødegå CNO-dosering og potentiel stress induceret af vores eksperimentelle design blev dyr udsat for en af fire betingelser: (1) unhabituated og injiceret med 10 mg/kg CNO; (2) habituated og injiceret med 10 mg/kg CNO; (3) unhabituated og injiceret med 1 mg/kg CNO; eller (4) habituated og injiceret med 1 mg/kg CNO. Tilvænning bestod af en 5-dages proces, der medførte håndtering, rektal temperatursonde, saltvandsinjektion og eksponering for plethysmografikammeret hver dag i 30 minutter, mens na-dyr kun blev udsat for en akklimatiseringsperiode på 20-45 minutter umiddelbart før dataindsamling som gjort i tidligere undersøgelser (Ray et al., 2011, 2013; Brust et al., 2014; Hennessy et al., 2017; Sun and Ray, 2017; Sun et al., 2017). Målte åndedrætsparametre inkluderede respirationsfrekvens RR, tidevandsvolumen (VT), minutventilation (VE), iltforbrug (VO2), minutventilation normaliseret til iltforbrug (VE/VO2), apnø frekvens, sukfrekvens og variationskoefficienter for interbreath interval og amplitude (periodisk og volumeninstabilitet). Som en ekstra kontrol sammenlignede vi også habituated og unhabituated vildtype dyr injiceret med saltvand.
CNO-hM4D forstyrrelse af Dbh-Cre neuroner resulterer i et hyperkapnisk underskud
under tre af betingelserne, DBH-Cre; RC::P_hm4d-dyr viste en reduktion i VE og VE/VO2 efter CNO – administration, mens søskendekontrol viste ingen forskelle i præ-og post-CNO-værdier. Uhabituerede dyr injiceret med 10 mg / kg CNO viste en signifikant reduceret RR (-12, 17%, p = 0, 034) og VT (-30, 87%, p = 0, 0016), hvilket resulterede i en reduktion i VE (-38, 64%, p = 0, 0031) og en lille reduktion i VO2 (-14, 25%, p = 0, 042) (figur 2). Reduktionen i VE var større end faldet i VO2, hvilket resulterede i en samlet reduktion i VE/VO2 (-26,89%, p = 0,0095). Uhabituerede dyr injiceret med 1 mg / kg CNO viste en tendens mod reduceret RR (-12,88%, p = 0,066) og signifikant reduceret VT (-16,52%, p = 0,00085) og VE (-28,08%, p = 0,0070), hvilket førte til en samlet reduktion i VE/VO2 (-22,23%, p = 0,016) (figur 3). Endelig viste habituerede dyr injiceret med 1 mg/kg CNO en reduceret RR (-10,77%, p = 0,074), tendens mod reduceret VE (-25,07%, p = 0,074) og reduceret samlet VE/VO2 (-23,70%, p = 0,024) (figur 4). Ingen signifikante ændringer i apnø frekvens, suk frekvens, periodisk eller volumen ustabilitet, eller temperatur blev set i nogen kohorte.
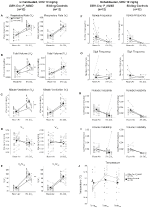
figur 2. Unhabituated DBH-Cre; RC:: P_hM4D dyr administreret 10 mg/kg CNO viser et hyperkapnisk underskud, mens søskendekontrol ikke viser nogen ændring. Efter CNO administration, DBH-Cre; RC::P_hm4d-dyr viser ingen ændring i rumluftventilation og reduktioner i RR (a), VT (B), VE (C), VO2 (D) og VE/VO2 (E) under hyperkapniske forhold uden ændringer i apnø frekvens (F), sukfrekvens (G), periodisk (H) eller volumeninstabilitet (I) og temperatur (J). Søskendekontrol viste ingen forskel i alle præ – og post-CNO-værdier.
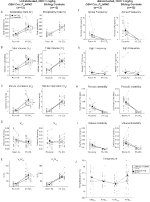
figur 3. Unhabituated DBH-Cre; RC:: P_hM4D dyr administreret 1 mg/kg CNO viser et hyperkapnisk underskud, mens søskendekontrol ikke viser nogen ændring. Efter CNO administration, DBH-Cre; RC::P_hm4d-dyr viser ingen signifikant ændring i rumluftventilation eller RR (A) og viste reduktioner i VT (B), VE (C) og VE/VO2 (E) under hyperkapniske forhold uden ændringer i VO2 (D), apnø frekvens (F), sukfrekvens (G), periodisk (H) eller volumeninstabilitet (I) og temperatur (J). Søskendekontrol viste ingen forskel i alle præ – og post-CNO-værdier.
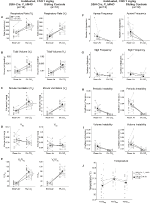
figur 4. Habituated DBH-Cre; RC:: P_hM4D dyr administreret 1 mg/kg CNO viser et hyperkapnisk underskud, mens søskendekontrol ikke viser nogen ændring. Efter CNO administration, DBH-Cre; RC:: p_hm4d-dyr viser ingen ændring i rumluftventilation og en reduktion i RR (A) og VE/VO2 (E) under hyperkapniske forhold uden signifikante ændringer i VT (B),VE (C),VO2 (D), apnø frekvens (F), sukfrekvens (G), periodisk (H) eller volumeninstabilitet (I) og temperatur (J). Søskendekontrol viste ingen forskel i alle præ – og post-CNO-værdier.
Habituerede Søskendekontrol injiceret med 10 mg / kg CNO viste et hyperkapnisk Ventilationsunderskud
i begge DBH-Cre; RC::P_hm4d og søskende kontrol habituerede dyr injiceret med 10 mg/kg CNO, bemærkede vi en signifikant reduktion I VE/VO2 (p = 0,0235) medieret af fald i RR (p = 0,00036) og VE (p = 0,037) (figur 5). I modsætning til de andre kohorter var der imidlertid ingen forskel mellem DBH-Cre; RC::P_hM4D og søskendekontroldyr i disse parametre: RR (-12,72 vs. -10,22%, p = 0,6268), VE (-24,88 vs. -23,94%, p = 0,4150) eller VE/VO2 (-15,4 vs. -22,55%, p = 0,4643).
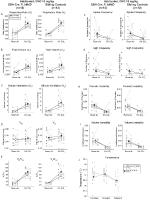
figur 5. Begge habituated DBH-Cre; RC::P_hm4d dyr og søskendekontrol administreret 10 mg/kg CNO viser et hyperkapnisk underskud. Efter CNO-administration viser DBH-Cre; RC::P_hM4D-dyr og søskendekontrol ingen ændring i rumluftventilation og en reduktion i RR (a), VE (C) og VE/VO2 (E) under hyperkapniske forhold uden signifikante ændringer i VT (B), VO2 (D), apnø frekvens (F), sukfrekvens (G), periodisk (H) eller volumeninstabilitet (I) og temperatur (J).
Habituated og Unhabituated vildtype Dyr injiceret med saltvand viste ingen ændringer Presalin og Postsaline
selvom der ikke blev observeret fænotyper i kontroller injiceret med 1 mg/kg CNO, behandlede vi muligheden for, at selve injektionen forårsagede den fænotype, der blev observeret i kontroller injiceret med 10 mg/kg CNO ved at teste habituated og unhabituated vildtype dyr injiceret med saltvand (figur 6). I både habituated og unhabituated kohorter viste dyr ingen forskel i respiratoriske parametre presalin og postsaline administration. Ingen signifikante ændringer i apnø frekvens, suk frekvens, periodisk eller volumen ustabilitet, eller temperatur blev set.
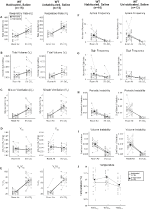
figur 6. Både uhabituerede og habituerede vildtype dyr administreret saltvand viser ingen forskel mellem værdier før og efter injektion. Efter saltopløsning viste både uhabiterede og habituerede vilde dyr ingen ændringer under rumluft eller hyperkapniske forhold i RR (a), VT (B), VE (C), VO2 (D), VE/VO2 (E), apnø frekvens (F), sukfrekvens (G), periodisk (H) eller volumeninstabilitet (I) og temperatur (J).
koncentrationsforhold mellem Closapin og CNO er højere i hjernen end i Serum
for at bestemme biotilgængeligheden af CNO og closapin målte vi deres koncentrationer i serum og hjerne via massespektrometri. Tredive minutter efter en intraperitoneal injektion af CNO hos mus findes CNO i lavere relativ overflod sammenlignet med dets rygmetabolit closapin i serum og hjerne for alle testede doser, 0,1 mg/kg (serum p = 0,0054, hjerne p = 0,0001) (figur 7A), 1 mg/kg (serum p > 0,05, hjerne p = 0,0197) (figur 7B) og 10 mg/kg (serum p = 0, 0036, hjerne p = 0.0005) (figur 7c). Når de samlede forhold mellem closapin og CNO blev analyseret, var de altid over nul og var højere i hjernen end i serum (vehikel p > 0,5, 0,1 mg/kg p > 0,5, 1 mg/kg p = 0,0018, 10 mg/kg p = 0,0160) (figur 7d).
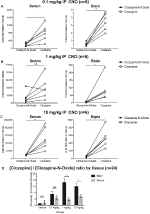
Figur 7. Forholdet mellem closapin og CNO er højere i hjernen end i serum. Serum-og hjerneprøver blev opsamlet fra vildtype mus 30 minutter efter CNO-administration i 3 forskellige doser 0,1 mg/kg (A), 1 mg/kg (B), 10 mg/kg (C). Individuelle datapunkter repræsenterer den relative overflod af closapin eller closapin opnået ved LC-MS. Closapin til CNO-forholdet for hver dosis, og SEM er også vist (d).
Diskussion
det oprindelige mål med denne undersøgelse var at undersøge rollen som DBH-Cre neuroner i respiratorisk fysiologi efter akut forstyrrelse hos uhæmmede og bevidste voksne dyr. Fordi noradrenalin og NA-neuroner spiller en veldokumenteret rolle i stressadfærd (Valentino og Van Bockstaele, 2008; Chen et al., 2018), forsøgte vi også at undersøge, om tilvænning til en sandsynlig stressende fysiologisk protokol ville have en effekt på de tidligere observerede respiratoriske fænotyper. I hele kroppen plethysmografi DREADD protokol, der anvendes i vores laboratorium og andre, håndteres dyr, rektalt probed for temperatur, udsat for et nyt miljø (plethysmography chamber) og intraperitonealt injiceret. Tidligere undersøgelser har vist, at håndtering og tilvænning til injektion og andre “rutinemæssige” procedurer kan ændre adfærdsmæssige og fysiologiske parametre, herunder respiration (Misslin et al., 1982; 1993; Lapin, 1995; Ryabinin et al., 1999). Andre anvendte stressfaktorer ændrer også respiration under både baseline og hyperkapniske ventilationsbetingelser (Isom og Elshovihy, 1982; Kinkead et al., 2001).
i vores undersøgelser, der anvendte høje doser CNO, fandt vi, at hM4D-medieret hæmning af DBH-Cre-definerede NA-neuroner resulterede i en reduceret hyperkapnisk refleks I VE/VO2 i fire definerede eksperimentelle kohorter med reduceret RR, VT og VE, der understøtter tidligere undersøgelser (Biancardi et al., 2008; Viemari, 2008; Gargaglioni et al., 2010). Imidlertid viste habituerede søskendekontrol, der fik en dosis på 10 mg/kg CNO anvendt i tidligere åndedrætsundersøgelser, et ventilationsunderskud under hyperkapniske forhold af samme størrelse som det, der ses hos DBH-Cre; RC::P_hM4D dyr. Ingen andre søskendekontrolgrupper viste denne fænotype, inklusive den habituerede kohorte, der kun modtog (1 mg/kg CNO) eller saltvand. Disse resultater antyder, at højere CNO-doser (10 mg/kg) har en effekt på det hyperkapniske respons, der er afmasket efter omfattende tilvænning, mens det formodentlig reducerer dyrestressniveauer, og at lavere CNO-doser ikke har en effekt på respiratorisk kontrol hos habituerede dyr. Disse resultater er også i overensstemmelse med arbejde fra Korsak et al. hvem demonstrerede, at CNO med lav dosis (2 mg/kg) ikke producerer effekter uden for målet i arbejdskapacitet i et træningsassay, der omfattede forudgående træning (Korsak et al., 2018) og Fleury Curado et al. hvem viste ingen lavdosis CNO (1 mg/kg) specifikke virkninger på respiratorisk output (Fleury Curado et al., 2018).
de øgede forhold mellem closapin og CNO-niveauer i serum og i hjernen (Figur 7) er i overensstemmelse med nylige undersøgelser, der antyder, at CNO let metaboliseres tilbage til closapin og viser større hjernepermeabilitet sammenlignet med CNO hos mus og andre steder (Jann et al., 1994; Chang et al., 1998; Guettier et al., 2009; Gomes et al., 2017; Raper et al., 2017). Det er imidlertid ikke klart, om de observerede off-target-effekter skyldes CNO eller closapin. Da vores hyperkapniske målinger fandt sted < 30 min efter CNO-applikation, er det sandsynligt, at off-target respiratoriske virkninger medieres af closapin. Vores resultater (Figur 7) viser høje relative niveauer af closapin i hjernen, selvom CNO ikke er helt fraværende. Imidlertid brugte Huckstepp og kolleger direkte CNO-applikation til den ventrale medulla hos bedøvede rotter for at demonstrere, at kun ved rumluft og ikke under hyperkapniske eller hypoksiske udfordringer har CNO-applikation en lille effekt, stigende frekvens og faldende ekspiratorisk varighed, men efterlader VT uændret uden nogen klar effekt observeret under hypercapnia (Huckstepp et al., 2015). I betragtning af den direkte anvendelse i bedøvede rotter og tidsramme for eksperimenter er det sandsynligt, at de små off-target-effekter, der blev set, blev medieret af CNO og ikke af closapin.
rygmetabolitten closapin er et almindeligt anvendt beroligende og antipsykotisk lægemiddel i schisofreni med mange endogene mål, herunder lav-høj affinitetsantagonistiske handlinger ved D1, D2 og D4 dopaminerge receptorer, 5-HT2A, 5-HT2C, 5-HT3, 5-HT6 og 5-HT7 serotonergiske receptorer, H1 histaminreceptorer og blandt andet Purpur-1 og Purpur-2 adrenerge receptorer (Fitton and Heel, 1990; Ashby 1996). De off-target-effekter, der ses her, kan skyldes forskellige eller kombinerede mekanismer og mål. Closapin kan påvirke respiration som et beroligende middel. En tidligere undersøgelse viste reduceret RR og VT under 5 og 10% CO2 under både langsom bølge-søvn og hurtig øjenbevægelse søvntilstande sammenlignet med den stille vågne tilstand hos mus (Nakamura et al., 2007). Alternativt kan hæmning af målrettede dreadd-ekspressive neuroner resultere i en antianksiogen eller angstdæmpende virkning svarende til vores tilvænningsprotokol for at afsløre CNO/closapin-medierede kemosensoriske effekter uden for målet. Begge forklaringer understøttes af flere undersøgelser, der har vist, at CO2 spiller en rolle i medfødte og lærte frygtresponser og angstrelateret adfærd., 2009; Feinstein et al., 2013; Taugher et al., 2014; Dlouhy et al., 2015; Vinter et al., 2017). Således kan neuroner, der er målrettet i nogle af disse undersøgelser, faktisk spille en rolle i kørsel af angstogene adfærdsmæssige reaktioner snarere end fysiologiske kemosensoriske reflekser, da både katekolaminerge og serotonerge systemer er involveret i frygt/angstadfærd og kemosensorisk homeostase (Brust et al., 2014; Hennessy et al., 2017).
omvendt kan de kemosensoriske fænotyper observeret med høje CNO-niveauer være ægte, da vi var i stand til at rekapitulere det NA-medierede hyperkapniske underskud ved CNO-doser, en størrelsesorden lavere i habituerede mus, mens kontrolgrupper ikke viste nogen CNO-effekt. Imidlertid er fulde sammenligninger på tværs af tidligere undersøgelser vanskelige på grund af manglen i nogle tilfælde af rapporterede VO2 -, VT -, RR-og VE/VO2-data. For eksempel kan ændringer i kropstemperatur eller metabolisk hastighed også påvirke på en række måder på åndedræts-og kemosensorisk output, og at plethysmografkammertemperaturer var meget forskellige i nogle tilfælde (34 vs. 30 kg C i vores undersøgelser), hvilket påvirker det dynamiske område for den barometriske komponent i bølgeformen og dermed tidevandsvolumen. Især så vi hverken mærkbare ændringer i åndedrætsbølgeformegenskaber under nogen af vores tilstande eller akut hjerte-åndedrætsstop, der blev observeret i vores tidligere høje dosis, hele rhombomere-undersøgelser (Sun et al., 2017).
vores resultater viser for første gang, at CNO har en uventet effekt på den hyperkapniske kemosensoriske refleks, der er afmasket af omfattende habituation. Det er vigtigt, på trods af høje niveauer, at en CNO-off-target-effekt tidligere var blevet udelukket på grund af manglen på fænotype i søskendekontrol, men som vi viser bliver tydelig ved tilvænning. Vi tilbyder en off-target karakterisering af CNO i musemodelsystemet for at supplere undersøgelser i rotte og ikke-menneskelige primater. Resultaterne her rejser muligheden for, at yderligere CNO-medierede, off-target effekter på kredsløbene under undersøgelse eller til autonome eller homeostatiske kredsløb kan eksistere, men kan maskeres i andre kontrollerede forsøg. Det er vigtigt, at disse data afslører, at efterforskere bør stræbe efter at bruge de minimale doser af den aktiverende ligand, der er mulig i kombination med høje niveauer af tilvænning, og at de korrekte kontroller skal inkluderes i kemiske genetiske manipulationer for fuldt ud at værdsætte og fortolke eksperimentelle data.
materialer og metoder
etisk godkendelse
undersøgelser blev godkendt af Baylor College of Medicine Institutional Animal Care and Use Committee under protokol AN-6171.
avl, genetisk baggrund og vedligeholdelse af mus
vi opretholdt kolonier af alle vores heterosygøse musestammer ved backcrossing til C57BL/6J mus og homosygøse stammer af søskendekryds. Til histologieksperimenter blev DBH-Cremice parret med den homosygøse AI9-mus (Madisen et al., 2010) (007909). Til plethysmografiforsøg blev DBH-Cre-mus parret med homosygøs RC::P_hM4D (Ray et al., 2011) mus til at udlede dyr, hvor alle mus bar RC::P_hM4D allel. Søskendedyr, der ikke arvede Cre-allelen, blev brugt som kontroldyr til det Cre-positive afkom. Rosa26-specifikke primere til AI9 -, RC::P_hM4D-og RC::ePe-musene var 5′ – gcacttgctctcccaaagtc, 5′ – GGGCGTACTGGCATATGATOG 5′ – CTTTAAGCCTGCCAGAAGA og giver et 495 bp-bånd (målrettet) og 330 bp-bånd (vægt). Cre-specifikke primere for alle rhombomere Cre drivere var 5 ‘- atcgccatcttccagcaggcgcaccattgccc og 5 ‘ – GCATTTCTGGGGGATTGCTTA og gav et 550 bp-bånd, hvis det var positivt. Til LC-MS-eksperimenter blev C57BL / 6J-mus opnået fra Center of Comparative Medicine (CCM), Baylor College of Medicine.
histologi
fire til otte uger gamle DBH-Cre; AI9 voksne mus blev transkardielt perfunderet med 0,1 M phosphatbufret saltvand (PBS) og derefter med 4% paraformaldehyd (PFA) i PBS. Hjerner blev dissekeret ud og faldt fast i 2 timer i 4% PFA før en PBS skylning og ækvilibrering i 20% saccharose i PBS. Hjerner blev derefter opdelt i 30 liter sektioner, monteret på dias og mærket med immunofluorescerende antistoffer. Vi farvede natten over med anti-tyrosin-antistof for at identificere katekolaminerge neuroner (1:1.000, Millipore AB152) efterfulgt af 2 timer med æsel anti-kanin Cy3 sekundær (1: 500, Jackson 711-165-152) i 0,1% Triton-H i PBS (PBST) med 5% æsel serum. Billeder blev samlet på et opretstående epifluorescerende mikroskop.
elektrofysiologi
horisontale hjerneskiver indeholdende locus coeruleus (300 liter) blev skåret med et vibratom (Leica VT 1000s, Leica Microsystems, Buffalo Grove, IL) fra voksne DBH-CRE; RR2P; RC::ePe-mus (~1 måned gammel) i 4 kunstig cerebrospinalvæske (ACSF). Skiverne blev nedsænket i et kammer perfunderet med iltet ACSF (95% O2 og 5% CO2) indeholdende i mM: 124 NaCl, 2,0 KCl, 1,3 MgSO4, 2,5 CaCl2, 1,2 KH2PO4, 25 NaHCO3 og 10 glucose (2-3 ml/min). Helcelleoptagelser blev udført ved 30 liter C ved anvendelse af konventionelle patch-clamp teknikker og en MultiClamp 700B forstærker (Molecular Devices, Union City, CA). GFP-positive neuroner fra locus coeruleus blev visuelt identificeret og efterfølgende afbildet af infrarød differentialinterferens kontrastvideo på scenen af et opretstående mikroskop (Aksioskope FS2, Carl Seiss, Oberkochen, Tyskland). Patchpipetter (modstande 4-6 MU) blev fyldt med (i mM): 110 K-gluconat, 10 KCI, 10 HEPES, 10 Na2-phosphocreatin, 2 Mg3-ATP og 0,2 Na3-GTP; pH blev justeret til 7,2 og osmolaritet til 300 mOsm. Holdingspotentialet var -70 mV. CNO blev anvendt bad.
plethysmografi
plethysmografi på bevidste, uhæmmede mus blev udført som beskrevet på 6 – til 12 uger gamle voksne dyr (Ray et al., 2011). Habituerede mus blev udsat for en 5-dages habituationsprotokol med hver dag bestående af flere minutters håndtering, temperatur taget af rektal probe, intraperitoneal saltvandsinjektion og 30 min i plethysmografikammeret. Mus blev derefter testet inden for 1 uge efter den sidste habituationsdag. Uhabituerede mus blev ikke udsat for håndtering eller plethysmografkammeret. Alle mus var na pristve til CNO og bruges kun en gang.
på testdagen blev mus taget fra deres hjemmebur, vejet og rektal temperatur blev taget. Dyr blev derefter anbragt i et gennemstrømning, temperaturstyret (vand kappet ved 30 liter C) plethysmografi kammer og fik lov til at akklimatisere i mindst 20 minutter i rumluft (21% O2/79% N2) betingelser. Efter akklimatisering og måling under rumluft blev kammergassen skiftet til en hyperkapnisk blanding af 5% CO2/21% O2/74% N2 i 20 minutter. Kammergas blev derefter skiftet tilbage til rumluft i 20 minutter. Musene blev kort fjernet til rektal temperaturmåling og intra-peritoneal injektion af CNO (National Institute of Mental Health Chemical Synthesis and Drug Supply Program) opløst i saltvand (1 eller 0,1 mg/ml) til en effektiv koncentration på henholdsvis 10 eller 1 mg/kg. Dyret blev returneret til kammeret i yderligere 20 minutter rumluft, 20 minutter hypercapnia og 20 minutter rumluft. Dyret blev derefter fjernet fra kammeret, og rektal temperatur blev taget umiddelbart efter afslutningen af eksperimentet.
væskekromatografi-massespektrometri
fireogtyve vildtypemus, jævnt fordelt efter køn, blev vejet og behandlet med 10 mg/kg CNO, 1 mg/kg CNO, 0, 1 mg/kg CNO eller køretøj. Tredive minutter efter injektion blev blodprøver opsamlet via hjertepunktur og anbragt i BD-Mikrobeholdere. Prøver blev centrifugeret ved 4 liter C ved 13.500 omdr. / min. i bænkcentrifuge, og supernatanter blev opsamlet. Serum og hjerner blev holdt på -20 C indtil ekstraktion.
opløsningsmidler af HPLC-kvalitet vand, acetonitrilchloroform og methanol og massespektrometri-grad myresyre blev opnået fra Sigma-Aldrich (St. Louis, MO). Kalibreringsopløsning indeholdende flere kalibreringsmidler i en opløsning af acetonitril, trifluoreddikesyre og vand blev købt fra Agilent Technologies. Metabolitter og interne standarder, herunder N-acetyl asparaginsyre-d3, tryptophan-15n2, glutaminsyre-d5, thymin-d4, gibberellinsyre, trans-seatin, jasmonsyre, 15N anthranilsyre og testosteron-D3 blev købt fra Sigma-Aldrich (St. Louis, MO). Microtainer R SST TM blev opnået fra Becton Dickinson (Franklin Lakes, NJ).
ekstraktion bestod af 750 liter iskold methanol:vand (4:1) indeholdende 20 liter spikede interne standarder, der blev tilsat til hver hjerneprøve (50 mg) og kvalitetskontrol og derefter blev homogeniseret i 1 min hver. Derefter blev 750 liter 100% acetonitril indeholdende 20 liter spikede interne standarder tilsat til vaskeprøve (100 liter) og kvalitetskontrol og derefter sonikeret i 5 minutter. Alle prøver blev centrifugeret ved 5.000 omdr. / min. i 10 minutter ved 4 liter C. Den resulterende supernatant blev opsamlet, og 20 liter blev injiceret i LC-MS.
alle prøver blev analyseret ved hjælp af 6.490 tredobbelt kvadrupol massespektrometer (Agilent Technologies, Santa Clara, CA) koblet til HPLC-system (Agilent Technologies, Santa Clara, CA) ved multiple reaction monitoring (MRM). Der blev opnået ca.8-11 datapunkter pr. påvist metabolit. Der blev påvist metabolitter af closapin, CNO og norclosapin (N-desmethylclosapin). ESI positiv tilstand blev anvendt i metode. HPLC-søjlen blev FRIKENDTUPLC C18-søjlen (100 -, 1,8-og 2,1-mm -, 100-mm -. Milford, MA, USA) med en strømningshastighed på 0,5 ml/min.
dataindsamling og analyse
plethysmografi
plethysmografi trykændringer blev målt ved hjælp af en Validyne DP45 differenstrykstransducer og referencekammer og CD15 bærerdemodulator og registreret med LabChartPro i realtid. Bølgeformer blev analyseret offline ved hjælp af LabChartPro og brugerdefineret skrevet MATLAB-kode for at bestemme respirationsfrekvens (RR), tidevandsvolumen (VT) (Ray et al., 2011), minutventilation (VE) og mønsteranalyse. Åndedrætsbølgeformer blev samlet offline i perioder, hvor dyret var i ro, og aflæsninger var fri for bevægelsesartefakter. Mindst 1 min kumulative data indsamlet fra spor mindst 10 s lang fra den sidste 10 min af en given eksperimentel tilstand blev analyseret. O2 forbrug blev bestemt ved at sammenligne gassammensætningen mellem kalibrering i et tomt kammer og levende vejrtrækning ved hjælp af en AEI iltføler og analysator. Kammertemperaturen blev konstant overvåget ved hjælp af en Termoarbejder MicroThermo 2 og sonde og blev registreret med LabChartPro i realtid.
Poincar-målinger og suk-og apnø-frekvens blev bestemt ved hjælp af 1 min bevægelsesfrie spor fra hver vejrtrækningstilstand. Suk blev defineret som et åndedræt med amplitude på mindst dobbelt så stor som det gennemsnitlige åndedræt. Apnøer blev defineret som et interbreath interval (IBI) mindst dobbelt så stort som det gennemsnitlige IBI. Variationskoefficienten (CV) for IBI og amplitude blev også beregnet ud fra den samme 1-min sporkompilering af hver vejrtrækningstilstand (standardfejl IBI eller amplitude/gennemsnitlig IBI eller amplitude).
statistik
plethysmografi
resultater (RR, VT, VE, VO2, VE/VO2, antal apnøer og suk og CV ‘ er af IBI og amplitude) for rumluft og hyperkapniske data blev sammenlignet mellem DBH-Cre; RC::P_hM4D-kohorter og søskendekontrol ved hjælp af en lineær regressionsmodel med blandede effekter med dyretype (eksperimentel vs. kontrol) og CNO-administration (før vs. efter injektion) som faste effekter og Animal id som en tilfældig effekt. Temperaturdata blev sammenlignet ved anvendelse af en lineær regressionsmodel med blandede effekter med dyretype (eksperimentel vs. kontrol) som en fast effekt. En p < 0,05 blev brugt til at angive statistisk signifikans, og individuelle datapunkter, gennemsnit og standardfejl for gennemsnittet vises på alle diagrammer.
væskekromatografi-massespektrometri
det opnåede areal under toppen for hver prøve blev normaliseret ved den interne kontrol og derefter til køretøjet, før statistisk analyse blev udført. Uparret t-test blev brugt til at sammenligne den relative overflod af closapin og CNO i hvert væv efter koncentrationsgruppe.
Etikerklæring
alle eksperimenter blev godkendt af det institutionelle Udvalg for dyrepleje og brug af Baylor College of Medicine. Forsøgene er i overensstemmelse med nationale standarder for pleje og anvendelse af forsøgsdyr, der er fastsat af Association for Assessment and Accreditation of Laboratory Animal Care.
Forfatterbidrag
JS, FS-M, MC-M og RR udtænkt og designet eksperimenterne. JS, FS-M og PS udførte eksperimenterne og bidrog til at analysere dataene. JS, FS-M, VM, MC-M og RR skrev papiret.
finansiering
denne undersøgelse blev støttet af tilskud R01HL130249 og R01HL130249-S1 fra National Heart, Lung and Blood Institute; marts af Dimes Basil O ‘ Connor-prisen; og McNair Medical Institute.
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
vi takker Drs. Shaila K. Mani og Benjamin Arenkiel for nyttige diskussioner. Vi takker også Baylor College of Medicine Proteomics Kernefacilitet til udførelse af væskekromatografi-massespektrometri.
Amiel, J., Dubreuil, V., Ramanantsoa, N., Fortin, G., Gallego, J., Brunet, J.-F., et al. (2009). PHOKS2B i respiratorisk kontrol: lektioner fra medfødt centralt hypoventilationssyndrom og dets musemodeller. Respir. Physiol. Neurobiol. 168, 125–132. doi: 10.1016 / j.resp.2009.03.005
CrossRef Fuld tekst | Google Scholar
and fil, S. E. (1993). Håndtering Historie af rotter ændrer adfærdsmæssige virkninger af lægemidler i den forhøjede plus-labyrint test af angst. EUR. J. Pharmacol. 235, 109–112. doi: 10.1016/0014-2999(93)90827-5
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
i 2007 blev han en af de mest kendte i verden. Udvikler låsen, så den passer til nøglen for at skabe en familie af G-proteinkoblede receptorer, der aktiveres kraftigt af en inert ligand. Proc. Natl. Acad. Sci. USA 104, 5163-5168. doi: 10.1073 / pnas.0700293104
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Ashby, C. R. og Vang, R. Y. (1996). Farmakologiske virkninger af det atypiske antipsykotiske lægemiddel closapin: en gennemgang. Synapse 24, 349-394. doi: 10.1002 / (SICI)1098-2396(199612)24:4<349::hjælp-SYN5>3.0.CO; 2-D
PubMed abstrakt | CrossRef fuldtekst | Google Scholar
Bahia, C. M. C. D. S., og Pereira, J. S. (2015). Obstruktiv søvnapnø og neurodegenerative sygdomme: en tovejs relation. Dement. Neuropsykol. 9, 9–15. doi: 10.1590 / S1980-57642015DN91000003
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Biancardi, V., B. Kurtcego, K. C., Almeida, M. C. og Gargaglioni, L. H. (2008). Locus coeruleus noradrenerge neuroner og CO2 kører til vejrtrækning. Pflugers Arch. – EUR. J. Physiol. 455, 1119–1128. doi: 10.1007 / s00424-007-0338-8
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Brust, R. D., Corcoran, A. E., Richerson, G. B., Nattie, E. og Dymecki, S. M. (2014). Funktionel og udviklingsmæssig identifikation af en molekylær undertype af hjerne serotonerg neuron specialiseret til at regulere vejrtrækningsdynamik. Celle Rep. 9, 2152-2165. doi: 10.1016 / j. celrep.2014.11.027
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest populære måder at gøre det på. (1998). Hos patienter med kronisk nyresvigt er der en risiko for, at sygdommen udvikler sig. Prog. Neuro-Psykofarmakol. Biol. Psykiatri 22, 723-739. doi: 10.1016 / S0278-5846(98)00035-9
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Chen, Y., Smith, K. G., Et Al. (2018). Genetisk identifikation af en population af noradrenerge neuroner impliceret i dæmpning af stressrelaterede reaktioner. Mol. Psykiatri 1-16. doi: 10.1038 / s41380-018-0245-8
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kaasaki, H., Oya, H., Musa, C., Et Al. (2015). Åndedræt hæmmes, når anfald spredes til amygdala og ved amygdala stimulering. J. Neurosci. 35, 10281–10289. doi: 10.1523 / JNEUROSCI.0888-15. 2015
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest almindelige årsager til, at der er tale om en alvorlig sygdom. (2013). Frygt og panik hos mennesker med bilateral amygdala skade. Nat. Neurosci. 16, 270–272. doi: 10.1038 / nn.3323
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Fitton, A. og Heel, R. C. (1990). Closapin. En gennemgang af dets farmakologiske egenskaber og terapeutisk anvendelse i schisofreni. Stoffer 40, 722-747. doi: 10.2165/00003495-199040050-00007
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Fleury Curado, T. A., Pho, H., Dergacheva, O., Berger, S., Lee, R., Freire, C., Et Al. (2018). Lyddæmpning af hypoglossale motoneuroner fører til søvnforstyrret vejrtrækning i magre mus. Front. Neurol. 9:962. doi: 10.3389 / fneur.2018.00962
CrossRef Fuld Tekst | Google Scholar
Gargaglioni, L. H. og Putnam, R. K. (2010). Locus coeruleus og Central kemosensitivitet. Respir. Physiol. Neurobiol. 173, 264–273. doi: 10.1016 / j.resp.2010.04.024
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest almindelige måder at gøre det på. (2017). Chemogenetics afsløret: DREADD belægning og aktivering via konverteret closapin. Videnskab 357, 503-507. doi: 10.1126 / videnskab.aan2475
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Guettier, J. M. I., Li, J. H., Rosemond, E., Et Al. (2009). En kemisk-genetisk tilgang til undersøgelse af G-proteinregulering af betacellefunktion in vivo. Proc. Natl. Acad. Sci. USA 106, 19197-19202. doi: 10.1073 / pnas.0906593106
CrossRef Fuld tekst | Google Scholar
Hakim, F., Gosal, D. og Kheirandish-Gosal, L. (2012). Sympatiske og katekolaminerge ændringer i søvnapnø med særlig vægt på børn. Front. Neurol. 3:7. doi: 10.3389 / fneur.2012.00007
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Hennessy, M. L., Corcoran, A. E., Brust, R. D., Chang, Y., Nattie, E. E. og Dymecki, S. M. (2017). Aktivitet af Tachykinin1-udtrykker Pet1 raphe neuroner modulerer respiratorisk kemorefleks. J. Neurosci. 37, 1807–1819. doi: 10.1523 / JNEUROSCI.2316-16. 2016
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Huckstepp, R. T. R., K. P., Henderson, L. E. og Feldman, J. L. (2015). Rolle af parafaciale kerner i kontrol af vejrtrækning hos voksne rotter. J. Neurosci. 35, 1052–1067. doi: 10.1523 / JNEUROSCI.2953-14. 2015
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Ilg, A.-K., Enkel, T., Bartsch, D. og B. L. L., F. (2018). Adfærdsmæssige virkninger af akut systemisk lavdosis closapin hos vilde rotter: implikationer for brugen af DREADDs i adfærdsmæssig neurovidenskab. Front. Opfør dig ordentligt. Neurosci. 12:173. doi: 10.3389 / fnbeh.2018.00173
CrossRef Fuld tekst | Google Scholar
Isom, G. E. og Elshovihy, R. M. (1982). Interaktion mellem akut og kronisk stress med respiration: ændring af nalokson. Pharmacol. Biochem. Opfør dig ordentligt. 16, 599–603. doi: 10.1016/0091-3057(82)90422-1
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Jann, M. V., Lam, Y. V. og Chang, H. V. (1994). Hurtig dannelse af closapin hos marsvin og mennesker efter administration af closapin. Bue. Int. Pharmacodyn. Ther. 328, 243–250.
PubMed Abstrakt | Google Scholar
Joober, R. (2010). Et særligt, dårligt forstået og underudnyttet molekyle. J. Psykiatri Neurosci. 35, 147–149. doi: 10.1503 / jpn.100055
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Kinkead, R., Dupenloup, L., Valois, N. og Gulemetova, R. (2001). Stressinduceret dæmpning af det hyperkapniske ventilationsrespons hos vågne rotter. J. Appl. Physiol. 90, 1729–1735. doi: 10.1152 / jappl.2001.90.5.1729
PubMed Abstrakt | CrossRef Fuld Tekst | Google Scholar
Klein, D. F. (1993). Falske kvælningsalarmer, spontane panik og relaterede tilstande. En integrativ hypotese. Bue. Psykiatri 50, 306-317. doi: 10.1001 / archpsyc.1993.01820160076009
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Korsak, A., Sheikhbahaei, S., Machhada, A., Gourine, A. V. og Huckstepp, R. T. R. (2018). Parafaciale neurons rolle i kontrollen af vejrtrækning under træning. Sci. Rep. 8: 400. doi: 10.1038 / s41598-017-17412-fra
CrossRef Fuld tekst | Google Scholar
Lapin, I. P. (1995). Kun kontrol: effekt af håndtering, sham injektion og intraperitoneal injektion af saltvand på opførsel af mus i en forhøjet plus-labyrint. J. Pharmacol. Toksikol. Metoder 34, 73-77. doi: 10.1016/1056-8719 (95) 00025-D
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
A. M., Alfonsi, G. og Matturri, L. (2013). Patofysiologi af det humane locus coeruleus-kompleks i føtal/neonatal pludselig uforklarlig død. Neurol. Res. 35, 44-53. doi: 10.1179/1743132812y. 0000000108
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
det er en af de mest almindelige årsager til, at en person er i stand til at vælge en person, der er i stand til at vælge en person, der er i stand til at vælge en person, der er i stand til at vælge en person, der er i stand til at vælge en person, der er i stand til at vælge en person, der ikke er i stand til at gøre det. (2016). Administration producerer adfærdsmæssige virkninger i Long-evans rotter: implikationer for design af DREADD-eksperimenter. eNeuro 3, 1-14. doi: 10.1523 / ENEURO.0219-16. 2016
PubMed abstrakt | CrossRef Fuld tekst / Google Scholar
det er en af de mest populære måder at gøre det på. (2010). Et robust CRE-rapporterings-og karakteriseringssystem med høj kapacitet til hele musens hjerne. Nat. Neurosci. 13, 133–140. doi: 10.1038 / nn.2467
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Mahler, S. V. og Aston-Jones, G. (2018). CNO ondt? Overvejelser for brugen af dreadds i adfærdsmæssig neurovidenskab. Neuropsykofarmakologi 43, 934-936. doi: 10.1038 / npp.2017.299
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest populære og mest populære måder at gøre det på. (2018). Dette er en af de mest almindelige årsager til, at en person er i stand til at tage medicin, når han eller hun er i stand til at tage medicin. Sci. Rep. 8, 3840. doi: 10.1038 / s41598-018-22116-fra
CrossRef Fuld tekst | Google Scholar
(1982). Effekter af isolation, håndtering og nyhed på hypofysen-adrenal respons i musen. Psykoneuroendokrinologi 7, 217-221. doi: 10.1016/0306-4530(82)90015-4
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
(2007). Agilance tilstand-afhængig dæmpning af hypercapnic kemorefleks og overdreven søvnapnø i oreksin knockout mus. J. Appl. Physiol. 102, 241–248. doi: 10.1152 / japplphysiol.00679.2006
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Obonai, T., Yasuhara, M., Nakamura, T. og Takashima, S. (1998). Catecholaminneuroner ændring i hjernestammen hos ofre for pludselig spædbarnsdødssyndrom. Pædiatri 101, 285-288. doi: 10.1542 / peds.101.2.285
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Takashima, S. og Tada, H. (2003). Alfa2-adrenerge receptorsubtype ændringer i hjernestammen i pludselig spædbarnsdødssyndrom. Tidlig Brummen. Dev. 75 (Suppl.), S129-S138. doi: 10.1016 / j. patofys.2004.01.015
CrossRef Fuld tekst | Google Scholar
(2018). Dosisafhængig reduktion i kokaininduceret bevægelse hos rotter med selvadministration af kokain i anamnesen. Neurosci. LETT. 674, 132–135. doi: 10.1016 / j.neulet.2018.03.045
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Ramanantsoa, N., Hirsch, M.-R., Thoby-Brisson, M., Dubreuil, V., Bouvier, J., Ruffault, P.-L., et al. (2011). Vejrtrækning uden CO (2) kemosensitivitet i betingede Phoks2b mutanter. J. Neurosci. 31, 12880–12888. doi: 10.1523 / JNEUROSCI.1721-11.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
Raper, J., Morrison, R. D., Daniels, J. S., Howell, L., Bachevalier, J., Wichmann, T., et al. (2017). Metabolism and distribution of clozapine-N-oxide: implications for nonhuman primate chemogenetics. ACS Chem. Neurosci. 8, 1570–1576. doi: 10.1021/acschemneuro.7b00079
PubMed Abstract | CrossRef Full Text | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Soriano, L. P., Nattie, E. E., and Dymecki, S. M. (2013). Egr2-neuroner styrer det voksne respiratoriske respons på hypercapnia. Brain Res. 1511, 115-125. doi: 10.1016 / j. brainres.2012.12.017
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Ray, R. S., Corcoran, A. E., Brust, R. D., Kim, J. C., Richerson, G. B., Nattie, E., Et Al. (2011). Nedsat åndedræts-og kropstemperaturkontrol ved akut serotonerg neuroninhibering. Videnskab 333, 637-642. doi: 10.1126 / videnskab.1205295
PubMed Abstrakt | CrossRef Fuld Tekst | Google Scholar
Ryabinin, A. E., Vang, Y. M. Finn, D. A. (1999). Forskellige niveauer af FOS immunoreaktivitet efter gentagen håndtering og injektionsstress i to indavlede stammer af mus. Pharmacol. Biochem. Opfør dig ordentligt. 63, 143–151. doi: 10.1016 / S0091-3057(98)00239-1
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Snyder, B., Shell, B., Cunningham, J. T. og Cunningham, R. L. (2017). Kronisk intermitterende hypoksi inducerer oksidativ stress og betændelse i hjerneområder forbundet med neurodegeneration i det tidlige stadium. Physiol. Rep. 5, 1-13. doi: 10.14814 / phy2.13258
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Sun, J. J., Huang, T.-V., Neul, J. L. og Ray, R. S. (2017). Embryonale baghjerne mønstergener afgrænser forskellige kardio-respiratoriske og metaboliske homeostatiske populationer hos voksne. Sci. Rep. 7: 9117. doi: 10.1038 / s41598-017-08810-4
CrossRef fuldtekst / Google Scholar
Sun, J. J. og Ray, R. S. (2017). TG (Th-Cre)FI172Gsat (Th-Cre) definerer neuroner, der er nødvendige for fuld hyperkapniske og hypoksiske reflekser. Biol. Åbn 6, 1200-1208. doi: 10.1242 / bio.026823
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest populære måder at gøre det på. (2014). Sengekernen i stria terminalis er kritisk for angstrelateret adfærd fremkaldt af CO2 og acidose. J. Neurosci. 34, 10247–10255. doi: 10.1523 / JNEUROSCI.1680-14. 2014
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Valentino, R. J. og Van Bockstaele, E. (2008). Konvergent regulering af locus coeruleus-aktivitet som et adaptivt respons på stress. EUR. J. Pharmacol. 583, 194–203. doi: 10.1016 / j. ejphar.2007.11.062
CrossRef Fuld tekst | Google Scholar
Verbraecken, J. og McNicholas (2013). Åndedrætsmekanik og ventilationskontrol i overlapningssyndrom og fedme hypoventilation. Respir. 14:132. doi: 10.1186/1465-9921-14-132
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Viemari, J.-C. (2008). Noradrenerg modulering af det respiratoriske neurale netværk. Respir. Physiol. Neurobiol. 164, 123–130. doi: 10.1016 / j.resp.2008.06.016
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
vinter, A., Ahlbrand, R., Naik, D. og Sah, R. (2017). Differentiel adfærdsfølsomhed over for CO2-inhalation hos rotter. Neurovidenskab 346, 423-433. doi: 10.1016 / j. neurovidenskab.2017.01.003
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de bedste måder at gøre dette på. (2009). Amygdala er en kemosensor, der registrerer kulsyre og acidose for at fremkalde frygtadfærd. Cell 139, 1012–1021. doi: 10.1016/j.cell.2009.10.029
PubMed Abstract | CrossRef Full Text | Google Scholar