- introduktion
- materialer og metoder
- høst af oocytter og cRNA-mikroinjektion
- isolering af Membranfraktioner og Immunoblotting
- immunhistokemi
- Frysefrakturelektronmikroskopi
- parret-Oocytanalyse og kvantificering af kontaktområder
- hydrostatisk Trykimpulsassay
- Capratinkubation
- statistisk analyse
- resultater
- udtryk for Claudin-5 og Integration i H. laevis oocyt plasmamembran
- Patches af stramme knudepunkter er synlige i Claudin-5, der udtrykker oocytter
- parret Oocytassay til analyse af Claudin Trans-interaktion
- hydrostatisk Trykimpulsassay afslører Claudin-specifikt kryds af Oocytpar
- inkubation med Caprate
- Diskussion
- Claudiner bidrager til stærkere vedhæftningsegenskaber
- konklusion
- data Tilgængelighedserklæring
- Etikerklæring
- Forfatterbidrag
- finansiering
- interessekonflikt
- anerkendelser
- supplerende materiale
introduktion
tight junction-proteinfamilien er afgørende for cellefysiologi, da mangel eller svækkelse er forbundet med sygdomme og dysfunktion i mange organer og væv, som vist f.eks. i det indre øre., 2001; Florian et al., 2003), nyre (Konrad et al., 2006; J. P. et al., 2009), mave-tarmkanalen (Resnick et al., 2005; Amasheh et al., 2009), epidermis (Furuse et al., 2002; Tebbe et al., 2002), og hjernekapillærer (Nitta et al., 2003; Jørgen et al., 2003). Claudiner repræsenterer en transmembranproteinfamilie bestående af mindst 27 medlemmer (Mineta et al., 2011). Ud over deres fire transmembrane spiraldomæner indeholder de to ekstracellulære sløjfer (ECL1 og ECL2), en kort N-terminal og en C-terminal (Susuki et al., 2014). Specifikke claudin-ekspressionsmønstre bestemmer og afspejler den selektive permeabilitet af epithelia, og claudinproteinernes evne til at interagere i cis (inden for den samme membran) og i trans (mellem membranerne i den nærliggende celle) tillader dannelse af barriereformning og poredannende stramme forbindelsesstrenge (Van Itallie og Anderson, 2006).
Claudin-5 udtrykkes stærkt i kapillær endotel og dominerer det stramme kryds (TJ) af blod-hjernebarrieren (BBB), da udtrykket er > 100 gange højere sammenlignet med enhver anden claudin (Ohtsuki et al., 2007). Desuden udtrykkes det i en række epitelvæv, herunder lunge (Soini, 2011), eksokrine væv (Comper et al., 2009), tarm (Garcia-Hernandes et al., 2017) og urinveje (Koda et al., 2011). Claudin – 5 forårsager imidlertid en stærkere barriere i hjernekapillærer end i andre væv (Reinhold og Rittner, 2017), og dens funktion er nedsat ved neurodegenerative og neuroinflammatoriske lidelser (Greene et al., 2019). Derfor er claudin – 5 afgørende for at opretholde BBB. Men BBB er ikke kun beskyttende, det begrænser også de terapeutiske muligheder, da lægemidler hindres for at gennemsyre denne barriere.
Nitta et al. (2003) rapporterede, at BBB er mere permeabel for molekyler af 800 Da i størrelse i claudin-5 mangelfulde mus sammenlignet med vildtype mus (Nitta et al., 2003). Dette var i overensstemmelse med transfektionseksperimenter, der demonstrerede en tætningseffekt af claudin-5 i Caco-2-cellemonolag (Amasheh et al., 2005).
en anden vigtig barriereformende claudin er claudin-3, som er rapporteret at selektivt forsegle barrieren mod passage af ioner af enten ladning og uladede opløste stoffer., 2010). Det udtrykkes også i det endotel-stramme kryds af hjernekapillærer, og dets funktionelle tab observeres i faser af mikrofartøjsbetændelse, glioblastom og choroidpleksus hos patienter med multipel sklerose (Engelhardt et al., 2001; Jørgen et al., 2003).
barriereegenskaber kan ændres dynamisk, som f. eks., inkubation med natriumkaprat blev påvist hurtigt og reversibelt at nedsætte transepitelial elektrisk modstand i den humane tarmcellelinie HT-29/B6 (Krug et al., 2013). Natrium caprate åbner forbigående claudin – 5 indeholdende barrierer ved stramme kryds af epitel-og endotelceller (Del Vecchio et al., 2012).
dette indikerer, at claudin-5 er et lovende mål for lægemiddelafgivelsesforbedring i BBB.
i denne undersøgelse havde vi til formål at anvende det heterologe ekspressionssystem af Ksenopus laevis oocytter (Vitsthum et al., 2019) til analyse af claudin-5 og claudin-3 interaktion og pertubation. På grund af manglen på endogene Celle-Celle-kontakter muliggør dette enkeltcelleekspressionssystem analysen af specifikke claudiner uden indblanding af andre stramme forbindelsesproteiner.
materialer og metoder
høst af oocytter og cRNA-mikroinjektion
oocytter blev opsamlet fra voksne kvindelige afrikanske klofrøer ved kirurgisk laparotomi. For anastesi, 0.2% MS222 (ethyl-3-aminobensoatmethansulfonat, Sigma-Aldrich, Taufkirchen, Tyskland) blev anvendt som en badopløsning i 5-10 minutter ved 20 liter C. Når kirurgisk anasthesi var nået, blev der foretaget indsnit i hud-og mavemuskler, og æggestokkens masse blev eksterioriseret og ovarievæv fjernet. Ved stuetemperatur i 90 minutter i 1,5 mg/ml collagenase Fisher Bioreagenter BP2649-1 (Fisher Scientific, Tyskland) opløst i oocyt Ringer-opløsning (ORi) som beskrevet af Vitsthum et al. (2019). Follikulære celler blev fjernet ved inkubation i Ca2 + – fri ORi indeholdende (i mM): NaCl (90), KCl (1), EGTA (triethylenglycol diamin tetraeddikesyre) (1), 5 HEPES (5); pH 7,4 i 10 minutter på en mekanisk ryster med 50 omdr. / min. Oocytter af trin V og VI (>1000 liter) blev injiceret (Nanoliter 2010, Verdens præcisionsinstrumenter, Sarasota, FL, USA) med 1 ng cRNA, der koder for human claudin-5, claudin-3 eller RNase-frit vand som kontroller. Injektionsvolumen var 50,6 nl pr. Efter injektion blev oocytter inkuberet ved 16 liter C i ORi 3 dage til proteinekspression.
isolering af Membranfraktioner og Immunoblotting
ti injicerede oocytter blev samlet til vestlig blot-analyse og resuspenderet i 500-homogeniseringsbuffer indeholdende (i mM) MgCl2 (5), NaH2PO4 (5), EDTA (ethylendiamintetraeddikesyre) (1), saccharose (80) og Tris (Tris)aminomethan) (20); pH 7.4. Oocytekstrakter blev centrifugeret to gange ved 200 omdr. / min. i 10 minutter ved 4 kg C for at kassere celleaffald. Supernatanten blev centrifugeret ved 13.000 omdr. / min i 30 minutter ved 4 liter C for at pille cellemembranen som beskrevet af Leduc-Nadeau et al. (2007). Pellets blev resuspenderet i 80-homogeniseringsbuffer til homogenisering. Proteinkvantificering blev udført kolorimetrisk ved anvendelse af Pierce 600 nm Protein Assay Kit (Thermo Fisher Scientific, Hennigsdorf, Tyskland) ifølge producentens instruktion i en 96 brøndplade. Pladelæseren (PerkinElmer Enspire Multimode Plate Reader, Valtham, MA, USA) blev justeret til 562 nm, og bovint Serumalbuminstandard (Thermo Fisher Scientific, Hennigsdorf, Tyskland), der spænder fra 125 til 2000 liter/ml, blev anvendt til evaluering. Forud for immunoblotting blev prøver blandet med 4 liter laemmli-buffer (Bio-Rad Laboratories, Munich, Tyskland), indlæst på en 10% SDS polyacrylamidgel og elektroforeseret. Til proteinoverførsel blev PVDF-membraner anvendt og blokeret i 5% fedtfri tør mælk i Tris-bufret saltvand i 120 minutter. Proteiner blev påvist ved immunoblotting ved anvendelse af primære antistoffer hævet mod claudin-3 eller claudin-5 (invitrogen #35-2500, #34-1700, #34-1600, Life Technologies, Carlsbad, CA, USA).
peroksidase-konjugeret ged anti-kanin og anti-mus antistoffer (#7074, #7076 celle signalering teknologi, Danvers, MA, USA) blev anvendt til at binde til de primære antistoffer og derfor inkuberet i mindst 45 minutter ved stuetemperatur. Til påvisning blev klarhed vestlige ECL Blotting substrat (#1705061, Bio-Rad Laboratories GmbH, Munich, Tyskland) anvendt, og signaler blev visualiseret af et ChemiDoc MP-system (Bio-Rad Laboratories).
immunhistokemi
injicerede oocytter blev fikseret i 4% PFA (16% paraformaldehyd, E15700, Science Service, Munich, Tyskland) i 4 timer ved stuetemperatur efterfulgt af dehydreringsgradient fra 70% ethanol til ksylol (Carl Roth, Karlsruhe, Tyskland) inden for 48 timer. prøver blev indlejret i paraffin og tværsnits (5 liter) ved anvendelse af en Leica RM 2245 microtome (Leica Microsystems Heidelberg, Tyskland). Kort før immunhistokemisk behandling blev paraffin fjernet via ksylol til ethanolgradient. Ikke-specifikke bindingssteder blev blokeret under anvendelse af 5% gedeserum i fosfatbufret saltvand og inkuberet med de samme primære antistoffer som til immunoblotting. Prøver blev inkuberet med de sekundære antistoffer Aleksa Fluor – 488 goat anti-rabbit og Aleksa Fluor-594 goat anti-mouse (Life Technologies, Carlsbad, CA, USA) og undersøgt ved konfokal laserscanning immunofluorescensmikroskopi (LSM 710, Seiss, Oberkochen, Tyskland).
Frysefrakturelektronmikroskopi
Frysefrakturelektronmikroskopi blev udført som rapporteret for nylig (Greene et al., 2019). Til fiksering blev injicerede oocytter inkuberet i glutaraldehyd (2,5% i 0,1 M cacodylatbuffer) natten over ved 4 liter C. Efter vask med cacodylatbuffer blev oocytter forberedt til frysefrakturering. Prøverne blev kryobeskyttet i 30% glycerol og frosset i flydende nitrogen. Efter brud og skygge med platin og kulstof (BAF400D; Balsatorer, Liechtenstein) blev det resterende organiske materiale fjernet ved en natriumhypochloritvask. Oocytter blev analyseret i et transmissionselektronmikroskop (EM-10, Seiss, Oberkochen, Tyskland) og fotograferet med et digitalkamera (tr Larsndle GmbH). Morfometrisk analyse af de stramme forbindelsesstrenge blev udført ved en forstørrelse på 20.000 liter.
parret-Oocytanalyse og kvantificering af kontaktområder
Mannitol blev implementeret for at krympe de injicerede oocytter og tillade en mekanisk fjernelse af vitellinmembranen ved hjælp af pincet uden at beskadige plasmamembranen. 5-10 oocytter blev anbragt i en petriskål (35 mm diameter, Thermo Fisher, Henningsdorf, Tyskland, #153066) fyldt med ORi. Mannitol blev tilsat og opløst, indtil der var opnået hypertonisk krympning af cellerne (ca.400 mOsmol/l i 10 minutter). Efter manuel devitellinisering blev oocytter straks overført til en 24 brøndplade (1. 86 cm2 overfladeareal, TPP Techno plastprodukter, Trasadingen, Sverige, # 92024) indeholdende 2 ml ORi. I hver brønd blev to celler forsigtigt grupperet ved at skubbe dem sammen med en Pasteur-pipette (1 ml, Thermo Fisher, Henningsdorf, Tyskland, #PP88SB) og en pæresonde.
oocytpar af claudin-5-ekspression (cldn5 − cldn5), claudin-3-ekspression (cldn3 − cldn3), claudin-3 og claudin-5 Co − ekspression (cldn3,5 − cldn3,5) og kontrol oocytter (kontrol-kontrol) blev holdt sammen i op til 48 timer i ORi ved 16 liter C.
Lysfeltmikroskopi blev anvendt til kvantificering af kontaktområdet for klyngede oocytter efter 1, 24 og 48 h. billeder af Na-oocytparene i 24 brøndkulturretter blev taget på disse tidspunkter ved hjælp af et Leica dmi6000 B-mikroskop (Leica Microsystems, vådtslar, Tyskland). Diameteren af kontaktområdet blev målt ved hjælp af mikronskalaen (LAS-af 3.2.0). Kontaktområder anses for at være cirkulære, og kontaktområdet blev således beregnet ved hjælp af cirkelligningen a=kur R2.
hydrostatisk Trykimpulsassay
Vitellinmembraner blev mekanisk fjernet som beskrevet før, og oocytter grupperet analagøst til det parrede oocytassay. Derudover blev blandede oocytpar (control–cldn5 og control–cldn3) testet i hydrostatisk trykimpuls (HPI) assay. Efter 24 timers stabilisering blev der oprettet en defineret hydrostatisk impuls ved hjælp af en enkeltkanals elektronisk pipette (EE-300r, Eppendorf Research Pro, programversion 2.06.00, Hamborg, Tyskland).
oocytter blev opbevaret i 24 brøndplader indeholdende 2 ml ORi, og central positionering blev kontrolleret inden påføring af pipetteringsvolumenet 250 liter ORi. Dispenseringshastigheden blev ensartet indstillet til maksimal hastighed, svarende til en dispenseringshastighed på 0,9 s. endvidere blev vinklen (45 liter) og anvendelsesafstanden (1,3 cm) ensartet påført. Omgivende tryk, viskositeten af ORi og diameteren af pipettespidsåbningen blev holdt under konstante forhold. Bright field mikroskopi blev anvendt til kvantificering af kontaktområder 30 min efter det hydrostatiske tryk blev påført og sammenlignet med kontaktområder før påføring. Den eksperimentelle opsætning er beskrevet i Figur 1.
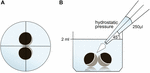
Figur 1. Eksperimentel opsætning af hydrostatisk trykimpulsassay (a) skematisk ovenfra af brønden: central positionering af oocytparet blev kontrolleret inden påføring af det hydrostatiske tryk. (B) skematisk sidebillede af brønden: 250 liter ORi blev tilsat ved hjælp af en elektronisk pipette med en enkelt kanal. Dispenseringshastigheden var ensartet indstillet til maksimal hastighed. Vinklen (45 liter) og påføringsafstanden blev anvendt ensartet. Omgivende tryk, viskositet af ORi og diameter af pipettespidsåbningen blev holdt under konstante forhold.
Capratinkubation
til capratinkubation blev natriumkaprat (#C4151, Sigma Aldrich, Taufkirchen, Tyskland) i endelige koncentrationer på 50, 100 og 500 liter eller ORi som referencegruppe føjet til oocytterne 24 timer efter parring. Oocytter blev holdt i 24 brøndplader indeholdende 2 ml ORi, og caprate-opløsning blev opløst i et defineret additionsvolumen på 250 liter ORi pr.brønd. Bredden af kontaktområdet blev kvantificeret ved 30, 60 og 120 minutter efter tilsætning.
statistisk analyse
statistisk analyse blev udført med JMP Pro 14.0.0 (NC, USA). Data præsenteres som medianer og vises som procentvis ændring baseret på den grupperede kombination ved de første undersøgelsespunkter. Figur 4, 5 præsenteres som Boksplotter, der viser den første kvartil (25 procent), medianen (50 procent) og den anden kvartil (75 procent). Knurhår trækkes ned til 10. percentil og op til 90.percentil. Normalfordelingen blev kontrolleret ved hjælp af Shapiro-test.
Kruskal–test blev brugt til flere sammenligninger efterfulgt af en Dunn-Bonferroni-korrektion. p-værdier er angivet som kontinuerlige tal.
resultater
udtryk for Claudin-5 og Integration i H. laevis oocyt plasmamembran
for at teste den vellykkede ekspression og integration af det stramme knudeprotein claudin-5 i oocytplasmamembranen, 3 dage efter injektion af claudin-5 cRNA, blev membranfraktioner analyseret ved immunoblotting. Alle prøver fra tre individuelle dyr (d1–d3) afslørede claudin-5-specifikke signaler ved 23 kDa, hvorimod Rnasefrie vandinjicerede oocytter ikke viste noget specifikt signal for claudin-5-ekspression (figur 2a).
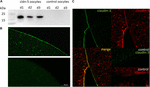
figur 2. Detektion af heterologt udtrykte claudiner i Ksenopus laevis oocytter. (A) Immunoblot analyse af tight junction protein claudin-5 i laevis oocytter af tre dyr (d1–d3). Cellemembranlysater blev udsat for SDS-side efterfulgt af immunoblot på PVDF-membraner. Membranerne blev inkuberet med primære antistoffer og sekundære peroksidase-konjugerede antistoffer (n = 3). (B) immunofluorescerende farvning afslørede specifikke claudin-5-signaler (grøn) i oocytmembraner af alle cRNA-injicerede oocytter, hvorimod der i vandinjekterede kontroller ikke blev påvist nogen claudin-specifikke signaler i konfokal mikroskopi. Repræsentative billeder af oocytter afledt af tre dyr. Skala barer: 50 liter. (C) immunofluorescerende farvning af claudin-5 og claudin-3, der udtrykker oocytter, afslørede specifikke claudin-3-signaler (grøn) og claudin-5-signaler (rød) i oocytmembraner af cRNA-injicerede oocytter, hvorimod der i vandinjekterede kontroller ikke blev påvist nogen claudin-specifikke signaler. Kolokalisering af udtrykte claudinproteiner i oocytplasmamembranen afsløres ved dobbelt immunofluorescerende farvning (gul). Skala bar: 10 liter.
til visualisering af de udtrykte proteiner i plasmamembranen blev immunhistokemiske farvestoffer udført og analyseret ved konfokal laserscanningsmikroskopi (figur 2b). Specifikke signaler blev detekteret og jævnt fordelt gennem plasmamembranen af claudin – 5, der udtrykker oocytter. I overensstemmelse med immunoblots blev der ikke påvist nogen specifikke signaler i kontrol-oocytplasmamembraner.
efter injektion af cRNA blev claudin-5 således udtrykt og integreret i plasmamembranen af H. laevis oocytter.
Co-ekspression af claudin-3 og claudin-5 i oocytpar afslørede specifikke signaler for claudin-5 (rød) og claudin-3 (grøn) i begge celler (figur 2C). Oocytplasmamembraner viste en fusion af de nærliggende celler tilvejebragt ved direkte cldn3,5–cldn3,5 interaktion (gul).
Patches af stramme knudepunkter er synlige i Claudin-5, der udtrykker oocytter
oocytplasmamembraner blev analyseret, og visualisering af stramme knudepunkter var vellykket (figur 3). Frysefrakturelektronmikroskopi viste pletter af strengmorfologi i plasmamembranerne af claudin-5 injicerede oocytter, og strengorganisation af claudin-5, der udtrykker oocytter, var meget organiseret og af vinkelform (figur 3a). Stramme forbindelsesstrenge blev primært detekteret i membranens protoplasmiske (P-) overflade. Claudin – 3 injicerede oocytter viste afrundede stærkt organiserede stramme krydsstrenge som tidligere rapporteret (Vitsthum et al., 2019; figur 3B). Frys frakturelektronmikroskopi af claudin – 3 og claudin-5 coudtrykkende oocytter afslørede fibriller, som begge bærer egenskaber af claudin-3 og claudin-5. Fibrilstrengarkitektur af coudtrykkende oocytter optrådte både afrundet og kompleks som claudin-3, der udtrykker oocytter, men også diskontinuerlige og mere vinklede som vist for claudin-5-ekspressive celler (figur 3c). Kontrol oocytter havde en typisk glat overflade (figur 3D).
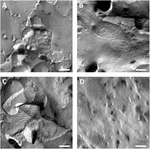
figur 3. Frys brudelektronmikroskopi. (A) Frysefrakturelektronmikroskopi afslører stramt knudeprotein cldn-5 som et netværk af kantede diskontinuerlige fibriller i rækker i Ksenopus laevis oocytter. (B) frysefrakturelektronmikroskopi afslører tæt knudeprotein cldn-3 som et net af afrundede fibriller i H. laevis oocytter. (C) Frysefrakturelektronmikroskopi af claudin-3 og claudin-5 coudtrykkende oocytter afslører fibriller, som begge bærer egenskaber af claudin-3 og claudin-5. (D) vand injiceret kontrol oocytter har en glat overflade. Repræsentative billeder af oocytter afledt af tre dyr. Vægtstang: 250 nm.
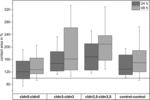
figur 4. Mediankontaktområder for klyngede oocytkombinationer cldn5-cldn5, cldn3–cldn3, cldn3,5–cldn3,5 og control-control 24 og 48 h efter klyngedannelse I % af de indledende kontaktområder kort efter klyngedannelse (n = 8-38).

figur 5. Kontaktområder for claudin-5, claudin-3 og coudtrykkende claudin-3 og claudin-5 oocytter i hydrostatisk trykimpuls (HPI) udfordring efter stabiliseringsperiode og 30 min efter HPI (n = 16-70, *p < 0,05, **p < 0.01, Kruskal-Vægis efterfulgt af en Dunn–Bonferroni korrektion).
parret Oocytassay til analyse af Claudin Trans-interaktion
alle grupperede kombinationer viste en tidsafhængig stigning i kontaktområdet over den målte tidsperiode (figur 4 og tabel 1).
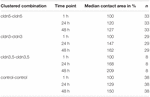
tabel 1. Oocytkontaktområder inden for 48 timer efter klyngedannelse.
kontaktområdet for vandinjekterede kontroloocytter steg til 129% efter 24 timer og 150% efter 48 timer. Grupperede par oocytter, der udtrykte claudin – 3, viste også en stigning i kontaktområder til 147% (24 timer) og 162% (48 timer). Grupperede par oocytter, der coudtrykkede claudin – 3 og claudin-5, viste kontaktområder på 168% (24 timer) og 209% (48 timer). Grupperede par oocytter, der udtrykte claudin-5 alene, viste kontaktområder på 120% (24 timer) og 127% (48 timer). Derfor var kontaktområderne i alle testede kombinationer sammenlignelige.
hydrostatisk Trykimpulsassay afslører Claudin-specifikt kryds af Oocytpar
i en separat tilgang blev oocytter, der udtrykker claudin-3 eller claudin-5 eller coudtrykker begge claudiner, grupperet efter mekanisk devitellinisering. Oocytter blev udfordret ved at anvende en HPI, og kontaktområder blev målt og beregnet 30 minutter efter udfordring og sammenlignet med indledende områder efter stabiliseringsperioden på 24 timer (figur 5 og tabel 2). Efter hydrostatisk trykudfordring faldt kontaktområdet for vandinjekterede kontroloocytter til 89%. Grupperede par af oocytter, der udtrykte claudin-5, claudin-3 eller coudtrykkende claudin-3 og claudin-5, bevarede større kontaktområder (97%, p = 0,0235; 96%, p = 0,003; 98%, p = 0,0253). Kontaktområderne for blandede vandinjicerede kontrol-oocytter og claudin-ekspressive oocytter (kontrol–cldn5 og kontrol–cldn3) adskiller sig ikke signifikant fra kontrol-oocytter (93%, p = 0,2900; 83%, p = 0,4455).
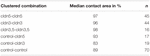
tabel 2. Hydrostatisk trykimpulsassay.
inkubation med Caprate
i et pilotinkubationseksperiment blev oocytter injiceret og parret i kombinationer, der enten udtrykte claudin-5 (cldn5–cldn5) eller injiceret med Rnasefrit vand som kontroller (kontrol–kontrol). Par blev inkuberet med endelige natriumkapratkoncentrationer på 50, 100 eller 500 liter. Inkubationen af oocytpar med ORi tjente som referencegruppe. Oocytter blev grupperet, og efter 24 timers stabilisering startede inkubation, og kontaktbredder blev målt 30, 60 og 120 minutter efter tilsætning (supplerende figur S1).
tilsætningen af ORi resulterede i et indledende fald i kontaktområder både i claudin-ekspression og vandinjicerede oocytpar, der er dispergeret 60 eller 120 minutter efter tilsætning. Dette er skitseret af den parabolske form af forbindelseslinjen mellem mediankontaktområderne over tid (røde kurver i supplerende figur S1). Men inkubation med 100 og 500 µM natrium caprate øget kontakt områder lidt (100 µM) eller kraftigt (500 µM) 30 min efter caprate ud fra 5,1 × 105 5.2 × 105 µm2 og 4,3 × 105 4,9 × 105 µm2.
Diskussion
i denne undersøgelse anvendte vi den klassiske model til transportører og human sygdomsmodellering (Tammaro et al., 2009; Nenni et al. Laevis oocytes, 2019), til en dybdegående analyse af claudin-5-interaktion og funktionelt Bidrag til krydsforseglingen. Til dette formål blev der etableret en ny tilgang, der introducerede en HPI til udfordrende interaktion inden for kontaktområdet for grupperede Ksenopus-oocytter.
Claudiner bidrager til stærkere vedhæftningsegenskaber
i overensstemmelse med tidligere resultater fra Vitsthum et al. (2019) førte enkeltklaudiner udtrykt i oocytter ikke til en stigning i interaktionskontaktområder sammenlignet med kontroloocytter. Imidlertid viste immunoblot og immunhistokemisk visualisering den vellykkede ekspression og integration i laevis oocytplasmamembran. Desuden tillod brugen af konfokal laserscanningsmikroskopi en præcis lokalisering af de udtrykte claudiner i plasmamembranen, da pinhole blokerede fluorescens uden for fokus. En kvantificering af immunhistokemiske signaler blev ikke forfulgt, da antistoffernes affinitet til binding af deres mål adskiller sig.
Vedula et al. (2009) brugte en mikropipette-aspirationsteknik til at undersøge aspekter af claudin-claudin-interaktion ved hjælp af L-fibroblaster transficeret med GFP-mærket occludin, cldn-1 og cldn-2. Den adskillelseskraft, der var nødvendig for at løsne to celler fra hinanden, var større i cldn-1 og cldn-2 transficerede celler (henholdsvis 2,8 og 2,3 nn). Selvom denne tilgang kan synes også lovende for claudin – claudin-interaktionsanalysen af at udtrykke oocytter, afslørede foreløbige tests, at en frigørelse af klyngede oocytter ikke er mulig uden forstyrrelse af oocytplasmamembranerne.
derfor blev forbindelsens kraft som en ny tilgang målt af en HPI. Selvom HPI ikke giver en kvantificering af separationskraften i absolutte værdier (f. eks., occludin, tricellulin, syltetøj-a). Claudiner bidrager til krydset mellem oocytpar, da de viser et større kontaktområde sammenlignet med vandindsprøjtede oocytter efter HPI. Dette indikerer stærk homofil transinteraktion mellem de claudin-ekspressive celler.
Strand fibril arkitektur er specifik for single claudins (Colegio et al., 2002). I frysefrakturelektronmikroskopi, claudin-3 blev rapporteret at samle et mere afrundet strengnetværkarbejde i løkkeformer i Ksenopus oocytter (Vitsthum et al., 2019; Figur 3b) mens claudin-5 i vores undersøgelse dannede et meshværk mere vinklet og bestilt i rækker. Billederne afslørede, at det stramme knudeprotein claudin – 5 danner et netværk af fibriller i diskontinuerlige, vinkelformede rækker i H. laevis oocytter. Dette er i overensstemmelse med claudin-5-tråde, der vides at forekomme som kæder af partikler forbundet med P-fasen (Piontek et al., 2011). I vores eksperimenter syntes geometrisk form af fibrillerne ikke at have nogen effekt på oocytternes vedhæftningsegenskaber. I overensstemmelse hermed blev paracellulær resistens rapporteret at være uafhængig af fibrilnummer og fibrildannende egenskaber (Colegio et al., 2002). Desuden er claudin – 3 og claudin-5 vist at have en lignende evne til homofil transinteraktion i HEK293 celler (Piontek et al., 2011).
denne model af laevis oocytter, der udtrykker enkelt tætte forbindelsesproteiner, tillader en observation af virkningen af stoffer som natrium caprate på dannelsen af kontaktområder mellem klyngede oocytter. Det kan derfor være et nyttigt værktøj til en tids-og omkostningseffektiv screening for stoffer, der påvirker den tætte forbindelsesbarriere.
Natriumkapratkoncentrationer på 100 og 500 liter kan have en beskyttende virkning på claudin-5, der udtrykker oocytter, hvilket resulterer i stigende kontaktområder efter 30 min inkubation.
Krug et al. (2013) demonstreret, at inkubation med natriumkaprat førte til et hurtigt og reversibelt fald i transepitelial resistens i human tarmcellelinje HT-29/B6. Desuden afslørede konfokal laserscanningsmikroskopi en markant reduktion af claudin-5 i HT-29/B6-celler behandlet med mellemkædede fedtsyrelaurat (Dittmann et al., 2014). Den første ekstracellulære sløjfe af claudiner (ECL1) er vigtig for barriereegenskaberne i det stramme kryds, mens den anden ekstracellulære sløjfe (ECL2) er involveret i strengdannelse af transinteraktion (Piontek et al., 2008; Rossa et al., 2014; Greene et al., 2019). Claudin – 5 bindemidler rettet mod ECL1 eller ECL2 af claudin-5 kan inducere intracellulær optagelse af det stramme knudeprotein og derved modvirke claudin-5 transinteraktioner i den stramme krydsforsegling mellem tilstødende celler og løsne det paracellulære rum (Hashimoto et al., 2017).
således antog vi, at inkubation med natriumkaprat ville føre til et fald i kontaktområdet af klyngede claudin-5, der udtrykker H. laevis oocytter. Uventet førte stigende koncentrationer af natriumkaprat (100 og 500 liter) til stigende kontaktområder af klyngede oocytter, der udtrykte claudin-5, hvilket indikerer en beskyttende virkning af natriumkaprat på den stramme krydsforsegling. Endvidere beskrives natriumkaprat for at inducere sammentrækning af den actomyosin perijunktionelle ring, hvilket udvider det paracellulære rum (Lindmark et al., 1998; Maher et al., 2009). Denne effekt er baseret på phosphorylering af den regulatoriske lette kæde af myosin via en phospholipase C-aktivering, som fører til en spaltning af phosphatidylinositol 4,5-bisphosphat (PIP2) i inositoltriphosphat (IP3) og diacylglycerol (Tomita et al., 1995). Det tætte forbindelseskompleks, der er forbundet med stilladsproteinet TSO-1 til actincytoskelettet, omfordeles derefter fra det stramme kryds til cytoplasmaet (Lindmark et al., 1998; Turner, 2000). En af de mest almindelige årsager til denne sygdom er, at det er en sygdom, der kan føre til en alvorlig sygdom., 1985). Selvom, før befrugtning, tjp-1–genekspression, genet, der koder for TSO-1, kun udtrykkes 1,9 TPM i oocytstadier V-VI (Session et al., 2016). En reduceret interaktion mellem claudinerne og det cytoskeletale stillads kan derfor forklare det uventede resultat af capratinkubationen. Er omfanget af fremtidige undersøgelser for at verificere det mekanistiske grundlag for fundet.
imidlertid blev en varianteffekt af natriumkaprat på den paracellulære permeabilitet beskrevet i litteraturen før. I Peyers Patchvæv taget fra tarmene hos voksne grise blev der påvist en lignende styrkende virkning. Claudin – 5 blev signifikant forøget efter inkubation med 5 mM caprate. I denne undersøgelse førte caprate til en signifikant højere transepitelial elektrisk modstand (TEER) i det follikelassocierede epitel (Radloff et al., 2019).
konklusion
i konklusion, heterolog ekspression af det stramme knudeprotein claudin – 5 i H. laevis oocytter tillader ny indsigt i enkelt claudins Bidrag til Celle-Celle interaktion og adhæsionsegenskaber af tilstødende celler. Laevis tight junction model for claudin – 5 tillader således analyse af BBB-komponenter i en enkeltcellemodel.
data Tilgængelighedserklæring
datasættene genereret til denne undersøgelse er tilgængelige på anmodning til den tilsvarende forfatter.
Etikerklæring
dyrebehandlingerne var i overensstemmelse med retningslinjerne i tysk lovgivning, med godkendelse af dyrevelfærdsofficer for Freie Universit Kript Berlin og under ledelse af Berlins Veterinærinspektorat (Landesamt f kurr Gesundheit og Sosiales Berlin, tilladelse G0025/16).
Forfatterbidrag
alle forfattere har læst og godkendt manuskriptet. NB og SA designede, planlagde og overvågede eksperimenterne og skrev manuskriptet. NB, LS, VC, RK og PF-B udførte forsøgene og dataanalysen.
finansiering
denne undersøgelse blev finansieret af Deutsche Forschungsgemeinschaft, Grant No. AM141/11-1 og Schaumann Stiftung.
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
vi takker Martin Grunau, Gisela Mans, Katharina s Larllig og Susanne Trappe for fremragende teknisk assistance. Vi anerkender støtte fra Open Access-Publikationsinitiativet fra Freie Universit Kript Berlin.
supplerende materiale
det supplerende materiale til denne artikel kan findes online på: https://www.frontiersin.org/articles/10.3389/fphys.2020.00857/full#supplementary-material
figur S1 / kontaktområder for claudin – 5, der udtrykker oocytpar i larm2 under inkubation med natriumkaprat i forskellige koncentrationer (henholdsvis n = 6-8) og vandindsprøjtede oocytter som kontroller (henholdsvis n = 5-7).
Amasheh, S., Dullat, S., Fromm, M., Schulke, J. D., Buhr, H. J. og Kroesen, A. J. (2009). Inflammet pose slimhinde besidder ændrede stramme kryds, der indikerer gentagelse af inflammatorisk tarmsygdom. Int. J. Colorect. Dis. 24, 1149–1156. doi: 10.1007 / s00384-009-0737-8
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Amasheh, S., Schmidt, T., Mahn, M., Florian, P., Mankert, J., Tavalali, S., et al. (2005). Bidrag fra claudin – 5 til barriereegenskaber i tætte kryds af epithelceller. Cellevæv Res. 321, 89-96. doi: 10.1007 / s00441-005-1101-0
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Colegio, O. R., Van Itallie, C., Rahner, C. og Anderson, J. M. (2002). Rollen af claudin ekstracellulære domæner i tæt kryds fibril arkitektur og paracellulær ladning selektivitet. Mol Biol Celle. 13:286.
Google Scholar
Comper, F., Antonello, D., Beghelli, S., Gobbo, S., Montagna, L., Pedersoli, P., et al. (2009). Ekspressionsmønster af claudins 5 og 7 skelner mellem solid-pseudopapillær fra pancreatoblastom, acinarcelle og endokrine tumorer i bugspytkirtlen. Er. J. Surg. Pathol. 33, 768–774. doi: 10.1097 / pas.0b013e3181957bc4
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
L., Tscheik, C., Tscheik, K., Helms, H. C., L., Blasig, R., et al. (2012). Natrium caprate åbner forbigående claudin-5-holdige barrierer ved stramme kryds af epitel-og endotelceller. Mol. Pharm. 9, 2523–2533. doi: 10.1021 / mp3001414
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Dittmann, I., Amasheh, M., Krug, S. M., Markov, A. G., Fromm, M. Og Amasheh, S. (2014). Laurat gennemsyrer den paracellulære vej for små molekyler i tarmepitelcellemodellen HT-29/B6 via åbning af de stramme kryds ved reversibel flytning af claudin-5. Pharm. Res. Dordr. 31, 2539–2548. doi: 10.1007 / s11095-014-1350-2
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
han er en af de mest kendte i verden, og han er en af de bedste i verden. (2001). Involvering af choroidpleksen i betændelse i centralnervesystemet. Microsc. Res. Teknik. 52, 112–129. doi: 10.1002/1097-0029(20010101)52:1<112::støtte-jemt13>3.0.co;2-5
CrossRef fuldtekst / Google Scholar
Florian, P., Amasheh, S., Lesidrensky, M., Todt, I., Bloedu, A., Ernst, A., et al. (2003). Claudiner i de stramme kryds af stria vaskularer marginale celler. Biochem. Biofys. Res. Co. 304, 5–10. doi: 10.1016 / s0006-291(03)00498-4
CrossRef fuldtekst / Google Scholar
Furuse, M., Hata, M., Furuse, K., Yoshida, Y., Haratake, A., Sugitani, Y., et al. (2002). Claudin-baserede stramme kryds er afgørende for pattedyrets epidermale barriere: en lektion fra claudin-1-mangelfulde mus. J. Cell Biol. 156, 1099–1111. doi: 10.1083 / jcb.200110122
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
(2017). Tarmepitel claudiner: ekspression og regulering i homeostase og inflammation. Ann. N. Y. Acad. Sci. 1397, 66–79. doi: 10.1111 / nyas.13360
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Greene, C., Hanley, N. og Campbell, M. (2019). Claudin-5: gatekeeper af neurologisk funktion. Væsker Barr. CNS 16: 4321.
Google Scholar
D., Amasheh, S., Pfaffenbach, S., Richter, J. F., Kausalya, P. J., Hunsiker, V., et al. (2009). Claudin – 16 påvirker transcellulær Cl – sekretion i mdck-celler. J. Physiol. Lond. 587, 3777–3793. doi: 10.1113 / jphysiol.2009.173401
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Hashimoto, Y., Shirakura, K., Okada, Y., Takeda, H., Endo, K., Tamura, M., et al. (2017). Claudin-5-bindemidler forbedrer permeationen af opløste stoffer over blod-hjernebarrieren i en pattedyrsmodel. J. Pharmacol. Eksp. Ther. 363, 275–283. doi: 10.1124 / jpet.117.243014
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Koda, R., L. N., Yaoita, E., Yoshida, Y., Tsukita, S., Tamura, A., et al. (2011). Ny ekspression af claudin-5 i glomerulære podocytter. Cellevæv Res. 343, 637-648. doi: 10.1007 / s00441-010-1117-y
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
det er en af de bedste måder at gøre dette på. (2006). Mutationer i det tætte knudegen claudin 19 (CLDN19) er forbundet med renal magnesiumudslip, nyresvigt og alvorlig okulær involvering. Er. J. Hum. Genet. 79, 949–957. doi: 10.1086/508617
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Krug, S. M., Amasheh, M., Dittmann, I., Christoffel, I., Fromm, M. Og Amasheh, S. (2013). Natrium caprate som en forstærker af makromolekylpermeation på tværs af tricellulære tætte kryds af tarmceller. Biomaterialer 34, 275-282. doi: 10.1016 / j.biomaterialer.2012.09.051
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Leduc-Nadeau, A., Lahjouji, K., Bissonnette, P., Lapointe, J. Y. og Bichet, D. G. (2007). Udarbejdelse af en ny teknik til oprensning af plasmamembraner fra Ksenopus laevis oocytter. Er. J. Physiol. Celle. 292, C1132-C1136.
Google Scholar
Lindmark, T., Kimura, Y. og Artursson, P. (1998). Absorptionsforbedring gennem intracellulær regulering af tæt krydspermeabilitet med mellemkædede fedtsyrer i Caco-2-celler. J. Pharmacol. Eksp. Ther. 284, 362–369.
Google Scholar
Maher, S., Leonard, T. V., Jacobsen, J. og Brayden, D. J. (2009). Sikkerhed og effekt af natrium caprate til fremme af oral lægemiddelabsorption: fra in vitro til klinikken. Adv. Drug Deliver. Åb 61, 1427-1449. doi: 10.1016 / j.addr.2009.09.006
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
det er en af de mest almindelige årsager til, at der ikke er nogen tvivl om, at der er tale om et problem. (2010). Claudin – 3 fungerer som en tætningskomponent i det stramme kryds for ioner af enten ladning og uladede opløste stoffer. Biochim. Biofys. Acta 1798, 2048-2057. doi: 10.1016 / j. bbamem.2010.07.014
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Mineta, K., Yamamoto, Y., Yamasaki, Y., Tanaka, H., Tada, Y., Saito, K., Et Al. (2011). Forudsagt udvidelse af claudin multigene-familien. FEBS Lett. 585, 606–612. doi: 10.1016 / j. febslet.2011.01.028
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Nenni, M. J., Fisher, M. E., James, C., Pells, T. J., Ponferrada, V., Chu, S., et al. (2019). Dette er en af de mest almindelige årsager til sygdommen hos mennesker. Front. Physiol. 10:154. doi: 10.3389 / fphys.2019.00154
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Nitta, T., Hata, M., Gotoh, S., Seo, Y., Sasaki, H., Hashimoto, N., Et Al. (2003). Størrelse-selektiv løsning af blod-hjernebarrieren i claudin-5-mangelfulde mus. J. Cell Biol. 161, 653–660. doi: 10.1083 / jcb.200302070
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Ohtsuki, S., Sato, S., Yamaguchi, H., Kamoi, M., Asashima, T. og Terasaki, T. (2007). Eksogen ekspression af claudin – 5 inducerer barriereegenskaber i dyrkede rotte hjernekapillære endotelceller. J. Cell Physiol. 210, 81–86. doi: 10.1002 / jcp.20823
PubMed Abstrakt / CrossRef Fuld Tekst / Google Scholar
Piontek, J., Fritsche, S., Cording, J., Richter, S., Hartvig, J., Valter, M., et al. (2011). Belysning af principperne for den molekylære organisation af heteropolymeriske stramme knudepunkter. Celle Mol. Livet Sci. 68, 3903–3918. doi: 10.1007 / s00018-011-0680-
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
det er en af de mest almindelige måder at gøre det på. (2008). Dannelse af tæt kryds: determinanter for homofil interaktion mellem klassiske claudiner. FASEB J. 22, 146-158. doi: 10.1096 / fj.07-8319com
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Radloff, J., Cornelius, V., Markov, A. G. og Amasheh, S. (2019). Caprate modulerer tarmbarrierefunktion i porcine Peyers patch follikelassocieret epitel. Int. J. Mol. Sci. 20:1418. doi: 10.3390 | Ijms20061418
PubMed abstrakt | CrossRef Fuld tekst / Google Scholar
Reinhold, A. K. og Rittner, H. L. (2017). Barrierefunktion i det perifere og centrale nervesystem-en gennemgang. Pflug. Bue. EUR. J. Phys. 469, 123–134. doi: 10.1007/s00424-016-1920-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Resnick, M. B., Gavilanez, M., Newton, E., Konkin, T., Bhattacharya, B., Britt, D. E., et al. (2005). Claudin expression in gastric adenocarcinomas: a tissue microarray study with prognostic correlation. Hum. Pathol. 36, 886–892. doi: 10.1016/j.humpath.2005.05.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Rossa, J., Ploeger, C., Vorreiter, F., Saleh, T., Protze, J., Gunzel, D., et al. (2014). Claudin – 3 og Claudin-5 proteinfoldning og samling i det stramme kryds styres af ikke-konserverede rester i segmenterne transmembran 3 (TM3) og ekstracellulær loop 2 (ECL2). J. Biol. Chem. 289, 7641–7653. doi: 10.1074 / jbc.m113. 531012
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
det er en af de bedste måder at gøre dette på. (2016). Genomudvikling i den allotetraploide frø Ksenopus laevis. Natur 538, 336-343.
Google Scholar
Soini, Y. (2011). Claudins i lungesygdomme. Hhv. Ress. 12: 70.
Google Scholar
T., Tani, K., Yamasaki, Y., Tamura, A., Ishitani, R., et al. (2014). Krystalstruktur af en claudin giver indsigt i arkitekturen af stramme kryds. Videnskab 344, 304-307. doi: 10.1126 / videnskab.1248571
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Tammaro, P., Shimomura, K. og Proks, P. (2009). “Ksenopus-oocytter som et heterologt ekspressionssystem til undersøgelse af ionkanaler med patch-clamp-teknikken” i kaliumkanaler: metoder og protokoller, Red. J. D. Lippiat, 127-139.
Google Scholar
C., Amasheh, S., Fromm, M., Assaf, C., Et Al. (2002). Tight junction proteiner: en ny klasse af integrerede membranproteiner-ekspression i human epidermis og i HaCaT keratinocytter. Arch Dermatol. Res. 294, 14-18. doi: 10.1007 / s00403-001-0290-y
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Tomita, M., Hayashi, M., S. (1995). Absorptionsforbedrende mekanisme af natrium caprate og decanoylcarnitin i Caco-2 celler. J. Pharmacol. Eksp. Ther. 272, 739–743.
Google Scholar
Turner, J. R. (2000). ‘At sætte klemmen’ på det stramme kryds: forståelse af cytoskeletal regulering. Sæd. Cell Dev. Biol. 11, 301–308. doi: 10.1006 / scdb.2000.0180
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
Van Itallie, C. M. og Anderson, J. M. (2006). Claudiner og epitelial paracellulær transport. Annu. Pastor Physiol. 68, 403–429. doi: 10.1146 / annurev.physiol.68.040104.131404
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Vedula, S. R. K., Lim, T. S., Kausalya, P. J., Lane, E. B., Rajagopal, G., Hunsiker, et al. (2009). Kvantificering af kræfter medieret af integrerede stramme knudeproteiner i Celle-Celle adhæsion. Eksper. Mech. 49, 3–9. doi: 10.1007/s11340-007-9113-1
CrossRef Full Text | Google Scholar
Vitzthum, C., Stein, L., Brunner, N., Knittel, R., Fallier-Becker, P., and Amasheh, S. (2019). Xenopus oocytes as a heterologous expression system for analysis of tight junction proteins. FASEB J. 33, 5312–5319. doi: 10.1096/fj.201801451rr
PubMed Abstract | CrossRef Full Text | Google Scholar
Wilcox, E. R., Burton, Q. L., Naz, S., Riazuddin, S., Smith, T. N., Ploplis, B., et al. (2001). Mutationer i genet, der koder for tæt kryds claudin-14, forårsager autosomal recessiv døvhed DFNB29. Celle 104, 165-172. doi: 10.1016 / s0092-8674(01)00200-8
CrossRef fuldtekst / Google Scholar
K., Kraus, J., Rascher-Eggstein, G., Liebner, S., Hamm, S., et al. (2003). Lokalisering af claudin – 3 i tætte kryds af blod-hjernebarrieren går selektivt tabt under eksperimentel autoimmun encephalomyelitis og humant Glioblastoma multiforme. Acta Neuropathol. 105, 586–592. doi: 10.1007 / s00401-003-0688-
PubMed abstrakt | CrossRef Fuld tekst | Google Scholar
(1985), C. C., Brun, D., Godsave, S. F., Kvarmby, J. og Heasman, J. Cytoskelet af Ksenopus oocytter og dets rolle i udviklingen. J. Embryol. Eksp. Morf. 89, 1–15.
Google Scholar