Betydning
der er en voksende interesse for circrna ‘ er på grund af deres implikation i mange biologiske processer og sygdomme ud over deres biomarkørpotentiale. De opdages hovedsageligt ved tilstedeværelsen af læsninger, der kortlægger deres backsplicing-kryds. Ikke desto mindre er circrna ‘er ikke længere de eneste udskrifter, der indeholder et sådant kryds, da nylige undersøgelser har afsløret, at cirkulære DNA’ er er almindelige og kan transkriberes, hvilket resulterer i udskrifter, der efterligner et circRNA-signal. Derfor kan denne nye type kimærisk transkription ændre den måde, hvorpå circRNA-analyse udføres, og påvirke nogle af de allerede rapporterede resultater.
er cirkulære RNA ‘ er de eneste Kimære udskrifter?
cirkulær RNA (circrna ‘ er) blev genopdaget for et par år siden som ikke-kanonisk splejsede RNA-former til stede i forskellige organismer, herunder mennesker., 2012; Jeck et al., 2013). De er kovalent lukkede transkripter dannet gennem en RNA-splejsningshændelse, hvor en splejsedonor af en nedstrøms ekson slutter sig til en opstrøms splejsningsacceptor, der fører til kovalent lukkede transkripter, der er kendetegnet ved tilstedeværelsen af et back-splejsningskryds, der gør circrna ‘ er skelnelige fra deres lineære modstykker (figur 1a) (Jhang et al., 2016; Helle, 2018).
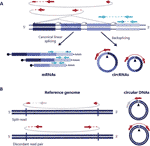
Figur 1 detektion af både cirkulært RNA og DNAs er baseret på aflæsninger, der spænder over krydset. (A) Backsplic sammenføjning mellem eksoner og kanonisk eksonforbindelse er afbildet med henholdsvis ikke-kontinuerlige og kontinuerlige linjer, der giver anledning til cirkrna ‘er og mRNA’ er. Parrede endelæsninger, der spænder over de bagspidsede kryds, vises i rødt, og parrede endelæsninger, der er i overensstemmelse med et lineært transkript, vises i blåt. (B) I lighed med circrna ‘er detekteres cirkulære DNA’ er (eccdna ‘ er) baseret på strukturelle læste varianter, der er i overensstemmelse med en cirkulationshændelse afbildet i rødt .
siden deres genopdagelse har det videnskabelige samfund henledt opmærksomheden på circrna ‘ er og har undersøgt deres involvering i flere cellulære processer inden for sundhed og sygdom (hake and Harries, 2017), deres potentielle rolle som biomarkører (Abu og Jamal, 2016) og deres regulatoriske funktioner (Floris et al., 2016). Circrna ‘ er er nu kendt for at være rigelige og stabile i cytosolen og kernen., 2012; Jeck et al., 2013; Li et al., 2015) og er også fundet Gratis i biofluider (Bahn et al., 2015; MC et al., 2015; Chen et al., 2018) og i ekstracellulære vesikler (Kyoung Mi et al., 2017). Biomarkørpotentialet for circrna ‘er er blevet intensivt undersøgt, faktisk er der blevet offentliggjort mange case-control-undersøgelser, der søger differentielt udtrykte circrna’ er, der kunne være biomarkører for forskellige sygdomme. Til dato har circrna ‘ er været impliceret i flere sygdomme, herunder kræft (Kristensen et al., 2017; Arnais et al., 2018), neurologiske lidelser (Akhter, 2018), hjerte-kar-sygdomme (Aufiero et al., 2019) og immunrelaterede sygdomme (Iparraguirre et al., 2017; Liu et al., 2019). På samme tid, at få fuldt ud at forstå deres biogenese, egenskaber, funktioner, og implikationer i menneskelig biologi forbliver som åbne spørgsmål for forskere på området.
selvom funktionen af de fleste circrna ‘er forbliver ukendt, er det vist, at nogle circrna’ er kan fungere som mikroRNA-svampe, der regulerer mikroRNA-niveauerne og deres aktivitet (Hansen et al., 2013; MC et al., 2013; Jens et al., 2016). De er involveret i genekspressionsregulering ved at regulere transkriptionen af deres forældregener og konkurrere med de lineære splejsnings-eller svampeproteiner., 2014; Li et al., 2018). Interessant nok har ribosomprofileringsundersøgelser for nylig vist, at circrna ‘ er også kan oversættes både in vitro og in vivo (Legnini et al., 2017; Yang et al., 2017).
hovedtræk ved circrna ‘ er, og ansvarlig for de fleste af deres særlige egenskaber, er deres cirkularitet. Derfor er det foruden at detektere deres karakteristiske back-splejsede kryds, at teste cirkulariteten af disse molekyler, et af nøglepunkterne i hver circRNA-undersøgelse. Ikke desto mindre har mange undersøgelser baseret deres opdagelse af circrna ‘er på total RNA og kan derved have fortolket nogle lineære kimære transkripter som circrna’ er, hvilket resulterer i falske positive circRNA-detektioner. For at omgå dette problem har de fleste undersøgelser bekræftet cirkulariteten af transkripterne fundet ved total RNA-sekv ved hjælp af RNase R, Northern blot eller elektroforetiske metoder (Jeck and Sharpless, 2014). Imidlertid har disse cirkulærhedsvalideringer også undertiden afsløret transkripter, der synes at være lineære, snarere end cirkulære, der bekræfter, at påvisning af cirkrna ‘ er startende fra total RNA kan føre til nogle falske positive. Disse falske positiver er blevet tilskrevet tekniske artefakter eller udskrifter afledt af usædvanlige begivenheder såsom ekson duplikationer eller transpliceringshændelser (Jeck and Sharpless, 2014; Sabo og Salsman, 2016). Når det er sagt, er muligheden for at have fundet ægte, biologisk aktive og funktionelle lineære transkripter, der indeholder en sekvens svarende til et backsplic-kryds (fra nu af kaldet kimære lineære transkripter), blevet overset noget, fordi en kilde til sådan lineær RNA ikke har været kendt for sunde celler.
cirkulære DNA ‘ er som kilde til Kimære lineære transkripter
det meste af det humane genom er organiseret i lineære kromosomer, men nogle undtagelser er længe blevet accepteret, såsom mitokondrie-DNA, og kromosomafvigelser såsom DNA-cirkler, der bærer onkogener (f.eks. dobbelt minutter) (Benner et al., 1991; Nathanson et al., 2014; Turner et al., 2017) og ringkromosomer (t., 2004). Det var først for nylig, at forskellige cirkulære DNA ‘er såsom mikrodna’ er (Shibata et al., 2012) eller ekstrachromosomale cirkulære DNA ‘er (eccdna’ er) viste sig også at stamme fra store dele af forskellige eukaryote genomer, herunder human og gær (m Larrller et al., 2015; Kumar et al., 2017; m Larsen et al., 2018).
cirkulære DNA ‘er dannes, når to ender af et lineært DNA er sammenføjet, hvilket resulterer i et kryds svarende til det backspliced kryds på circrna’ er, der almindeligvis kaldes breakpoint junction, der detekteres baseret på strukturelle læste varianter, der er i overensstemmelse med en cirkulationshændelse (Gresham et al., 2010; m Larsen et al., 2018; Prada-Luengo et al., 2019) (figur 1b). They usually range from a hundred bases to megabase circles and can contain full exons and genes (Shibata et al., 2012; Møller et al., 2015; Kumar et al., 2017; Turner et al., 2017; Møller et al., 2018) and while some regions of the genome are more commonly found on circular DNA (Sinclair and Guarente, 1997; ; Møller et al., 2016; Turner et al., 2017; Møller et al., 2018), most circular DNA appear to occur at random (Shibata et al., 2012; Møller et al., 2015; Kumar et al., 2017; Møller et al., 2018).
Interestingly, in a recent paper Møller et al. identificerede tusindvis af eccdna ‘ er i leukocytter og muskelceller i sunde kontroller. Med ideen om at undersøge, om eccdna ‘ er kunne transkriberes, blev et mRNA-Bibliotek også sekventeret fra muskelvæv og analyseret for transkriptionshændelser på tværs af breakpoint-krydset mellem det detekterede Eccdna, der fandt flere kampe (M., 2018). Dette fund antyder, at cirkrna ‘ers cirkulære DNA i sundt væv transkriberes, hvilket giver anledning til lineære og polyadenylerede transkripter, der vil bære en sekvens svarende til backsplic-sekvensen af circrna’ er (M-Rristller et al., 2018) (figur 2).
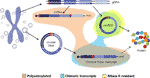
figur 2 grafisk repræsentation af forskellige udskrifter opstod fra enten genomisk DNA eller cirkulært DNA. Eksoner er farvet i lilla, og de bagspidsede kryds eller kimære kryds er vist med rødt. Polyadenylerede, kimære kryds indeholdende og RNase r resistente udskrifter er fremhævet i henholdsvis orange, blå og gul.
transkriptionsbeviset for cirkulære DNA ‘er, sammen med deres overflod, får os til at antyde, at cirkulære DNA’ er kan være en naturlig kilde til en betydelig mængde lineære RNA ‘ er, der bærer kimære kryds. I mange tilfælde, disse kimære kryds kan ikke skelnes fra backsplicing-kryds af circrna ‘ er, og derfor kan de være forvirrende faktorer i circRNA-undersøgelser. I de følgende afsnit vil vi forklare de data, der understøtter dette forslag.
circRNA detektion: Alt, hvad der glitter, er ikke guld
som tidligere introduceret dannes circrna ‘ er, selvom en ikke-kanonisk splejsningshændelse kaldes backsplicing. Transkripter, der er resultatet af denne backsplicing-begivenhed, har en kovalent lukket sløjfestruktur med hverken 5′-3′ polaritet eller en polyadenyleret hale, og endnu vigtigere er de kendetegnet ved tilstedeværelsen af en krypteret eksonordre i forhold til det lineære transkript., 2016; Helle, 2018). Denne krypterede eksonordre bliver tydelig i det backspliced kryds, der forbinder en 5′ nedstrøms sekvens med en opstrøms 3’ sekvens. Således udnytter alle circRNA-detektionsalgoritmerne tilstedeværelsen af de back-splejsede kryds som et diagnostisk træk til identifikation af circRNA (figur 1a).
forskellige metoder er blevet tilpasset til påvisning af disse back-splejsede kryds. Kommercielle arrays indeholdende prober, der er målrettet mod disse backspliced regioner, er blevet brugt i vid udstrækning i biomarkørscreeningsundersøgelser (Iparraguirre et al., 2017; Liu et al., 2017; Sui et al., 2017; Li et al., 2018). Den efterfølgende Validering er ofte også baseret på forstærkningen af backsplic-krydsene ved hjælp af divergerende primere (Panda og Gorospe, 2018). Mange andre papirer har udført en sekventeringsanalyse med høj kapacitet, der overvinder en af de vigtigste begrænsninger i arrays, der gør det muligt at opdage ikke kun de kommenterede circrna ‘ er, men også de novo RNA-cirkulariseringshændelser fra genomiske regioner, hvor ingen circRNA blev kommenteret af tidligere undersøgelser. Flere bioinformatiske rørledninger er udviklet til påvisning af cirkrna ‘ er i RNA-Sekv-datasæt, men alle er baseret på tilstedeværelsen af læsninger, der krydser back-splejsningskrydsene, og at finde den mest pålidelige er stadig en udfordring for bioinformatikere (Hansen et al., 2016; Hansen, 2018; Prada-Luengo et al., 2019).
to hovedmetoder kan følges til påvisning af circrna ‘ er i RNA-Sekv-data. For det første er mange circRNA RNA-sekv-undersøgelser baseret på RNase r-behandlede prøver for at nedbryde alle de lineære RNA ‘ er før sekventering. Selvom denne fremgangsmåde er specielt designet til circRNA-detektion, er det værd at bemærke, at RNase r-nedbrydning er variabel, at der er sjældne tilfælde af RNase R-resistente lineære RNA ‘er og RNase r-følsomme circrna’ er (2016), og at denne behandling kræver et højt RNA-input, som kan være begrænsende for nogle væv. Andre circRNA-undersøgelser vælger at sekvensere enten total, ribosomalt-udtømt (ribo -) eller ikke-polyadenyleret (polyA-) RNA, hvor både lineære og cirkulære RNA ‘ er kan findes., 2012; MC et al., 2013; Broadbent et al., 2015; Lu et al., 2015; Et al., 2015). Denne tilgang undgår brugen af RNase R, hvilket reducerer den RNA-mængde, der er nødvendig til sekventeringen, og gør det muligt at studere ekspressionen af andre typer RNA ‘ er fra det samme datasæt. Det er blevet påvist, at med en god sekventeringsdybde og kvalitet og en omhyggelig dataanalyse kan ægte circrna ‘ er detekteres fra total RNA-sekventering., 2017) er der imidlertid i denne anden tilgang behov for en senere cirkulærhedsbekræftelse for circrna ‘ er.
med opdagelsen af lineære kimære RNA ‘er transkriberet fra cirkulære DNA’ er er circrna ‘ er ikke længere de eneste udskrifter med kimære kryds. Derfor er det yderst vigtigt at bemærke, at mens den første tilgang væsentligt vil berige RNA-prøven i circrna ‘er, så de fleste af de detekterede kimære kryds svarer til ægte circrna’ er, kan den anden overvurdere antallet af circRNA-transkripter ved at tildele circrna ‘er signalet, der kommer fra både circrna’ er og de lineære kimære transkripter transkriberet fra cirkulære DNA ‘ er. Under hensyntagen til sameksistensen af cirkrna ‘er og lineære kimære transkripter får behovet for cirkularitetstest og funktionalitetsanalyser derfor betydning, og der bør udvises særlig omhu med hensyn til ikke kun eksperimentelle, men også beregningsmetoder for at undgå at forveksle kimære transkripter fra cirkulære DNA ‘er med cirkrna’ er dannet ved backsplicing.
Diskussion
circRNA-feltet er stadig på et tidligt stadium, men circrna ‘ er har allerede vist sig at være forbløffende molekyler, impliceret i mange processer, med et stort biomarkørpotentiale, og det kan også ændre den måde, vi forstår transkriptions-og oversættelsesprocesserne på. Af disse grunde får de opmærksomhed, og circRNA-feltet er i øjeblikket et af de mest aktive RNA-forskningsområder. Der er dog stadig mange konflikter, kontroverser og åbne spørgsmål (Li, 2019), der skal diskuteres.
i denne rapport og i lyset af de nylige fremskridt inden for det cirkulære DNA-felt ønsker vi at påpege transkriptionen fra ekstrachromosomalt cirkulært DNA som en af de vigtigste naturlige kilder til lineære transkriptioner med back-splejsede signaler, der kunne forstyrre circRNA-data (M., 2018). Fra nu af, bortset fra de tekniske artefakter, duplikationer og transpliceringshændelser, der kan føre til falske positiver i circRNA-detekteringen, bør vi også tage højde for eksistensen af denne nye type kimære udskrifter. Derfor er cirkulationstest og funktionelle analyser vigtigere end nogensinde.
under alle omstændigheder bør disse kimære lineære transkripter ikke kun betragtes som en simpel forvirrende faktor for circRNA-undersøgelser. På trods af de tekniske implikationer for circRNA-karakteriseringen tilføjer eksistensen af disse circRNA-lignende kimære lineære RNA-molekyler, der kommer fra eccdna ‘er, en ny type molekyle til den stadigt voksende liste over RNA’ er og udvider vores vision om transkriptomets kompleksitet og dets regulering. Desuden kunne disse lineære RNA-molekyler, der kommer fra eccdna, også præsentere funktioner svarende til circRNA, herunder regulatoriske funktioner eller potentialet til at blive oversat. Genprodukter fra eccdna-udskrifter kan potentielt bidrage til fænotypen af somatiske celler og væv som rapporteret i gær (Gresham et al., 2010; Demeke et al., 2015). Men i dette voksende felt er der behov for flere data og forskning for at begynde at ridse overfladen af isbjerget.
Forfatterbidrag
LI, IP-L, BR og DO skrev papiret.
finansiering
denne undersøgelse er finansieret af Instituto De Salud Carlos III gennem projektet “PI17/00189” (medfinansieret af Den Europæiske Fond for Regionaludvikling/Den Europæiske Socialfond) “investering i din fremtid”). IP-L og BR blev støttet af Det Frie Forskningsråd, 6108-00171b og LI blev støttet af Institut for uddannelse af den baskiske regering .
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.