inzeráty:
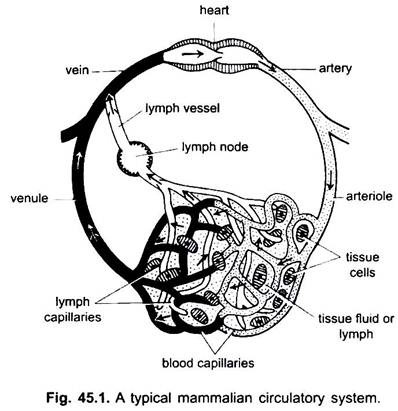
v oběhovém systému obratlovců existují dva systémy komplikovaně větvených trubek, které se rozvětvují po celém těle a přenášejí tekutiny do tkání. Jedná se o krevní cévní systém a lymfatický systém. Pokud se používá termín oběhový systém, vztahuje se pouze na krevní cévní systém.
krevní cévní systém je uzavřený systém u obratlovců; má kontraktilní srdce a kontinuální trubice zvané cévy. Lymfatický systém je otevřený systém s lymfatickými prostory. Krevní cévy, které odvádějí krev ze srdce, jsou tepny, které se dělí na tenčí arterioly a rozvětvují se na extrémně tenké a malé kapiláry.
stěna kapiláry je vyrobena z jedné vrstvy střapcovitých endotelových buněk. Každá krevní céva, včetně srdce, má výstelku endotelových buněk nebo endotelu. Krev nepřichází do styku přímo s tkáňovými buňkami.
inzeráty:
látky procházejí z A do kapilár tkáňovou tekutinou obsaženou v tkáňových prostorech mezi buňkami. Výměna látek mezi krví a tkáňovými buňkami těla probíhá přes kapilární stěny. To je způsobeno pinocytózou v endotelových buňkách kapilár.
pod elektronovým mikroskopem vykazují endotelové buňky mnoho malých váčků, které jsou invaginacemi plazmatických membrán. Tyto vezikuly se pohybují z jedné strany buňky a uvolňují jejich obsah na opačné straně.
krev tedy dává buňkám kyslík, živný materiál a hormony pro metabolismus a buňky vydávají vodu, oxid uhličitý a dusíkaté odpady do krve, aby je přenesly do vylučovacích orgánů pro jejich rychlou eliminaci.
kapiláry tvoří síť ve všech tělesných tkáních kromě chrupavky a epitelu. Z kapilár krev přechází do tenkých žilek, které se spojují a vytvářejí žíly, které přenášejí krev směrem k srdci. Ale některé žíly (portální žíly, renální žíly a jaterní žíly) mají kapiláry, které jsou stejně jako tepny. Ale veškerá krev neprochází kapilárami do žil.
inzeráty:
existují také některé kanály mezi arterioly a žilkami ležící v některých orgánech, jako je kůže. Tam jsou také arteriovenózní anastomózy mezi arterioly a venuly nalezené v číslicích. Funkce těchto spojení není jasně známa, i když se tvrdí, že regulují krevní tlak a oběh.
na některých místech dochází k výměně materiálu mezi tkáněmi a krví přes tenkostěnné prostory nebo sinusoidy. V některých orgánech může krevní céva tvořit stočenou síť drobných cév zvaných rete mirabile (ledviny, vzduchový močový měchýř).
části oběhového systému:
inzeráty:
u strun je oběhový systém uzavřeného typu. Tento typ se také vyskytuje u annelidů (Bezobratlých). Jiný typ je otevřeného typu, ve kterém nejsou nalezeny kapiláry. Vyskytuje se u měkkýšů a členovců. Zde krev protéká tepnami do různých orgánů, které procházejí krevními prostory nebo dutinami a pak znovu do cév (žil) do srdce.
oběhový systém zahrnuje srdce, tepny, žíly, kapiláry a krev. Srdce je modifikovaná krevní céva se svalovými stěnami, která se pravidelně stahuje, aby pumpovala krev do různých částí těla určitými cévami.
tepny a jejich větve tvoří arteriální systém, který nese krev ze srdce. Žíly a jejich přítoky tvoří žilní systém, který odebírá krev z kapilár tepen nebo arteriol a přenáší ji do srdce.
portálový systém:
v portálovém systému se krev nevrací přímo do srdce, ale v průběhu vracející se krve je vložený orgán (játra nebo ledviny). Žíla přinášející krev začíná v kapilárách a končí v kapilárách, dotyčná žíla působí jako aferentní i eferentní céva, aferentní cévy končí v kapilárách stejně jako tepny, pak se krev shromažďuje do systémových žil.
všichni obratlovci mají jaterní portální systém, ve kterém krev prochází do dvou sad kapilár v játrech. Nižší obratlovci a embrya vyšších obratlovců mají renální portální systém, ve kterém krev prochází dvěma sadami kapilár v ledvinách před dosažením srdce. Kapiláry v hypofýze tvoří hypofyzární portální systém, který je malý, ale důležitý systém.
lymfatický systém:
vyskytuje se v chordátech kromě cyklostomů a chrupavčitých ryb. Zahrnuje lymfatické a lymfatické cévy. Lymfa je tkáňová tekutina, která se nachází mezi tělními buňkami. Je to krevní plazma minus červené krvinky a některé bílkoviny.
lymfatické kapiláry tvořící síť tenkých slepých končících cév, které shromažďují lymfy. Lymfatické cévy jsou tenkostěnné cévy tvořené spojením krevních kapilár. Tyto prázdné do žil. Lymfatické uzliny se nacházejí na lymfatických cévách u savců. Ty tvoří lymfocyty krve používané k obraně těla proti nemocem.
vývoj srdce u obratlovců:
inzeráty:
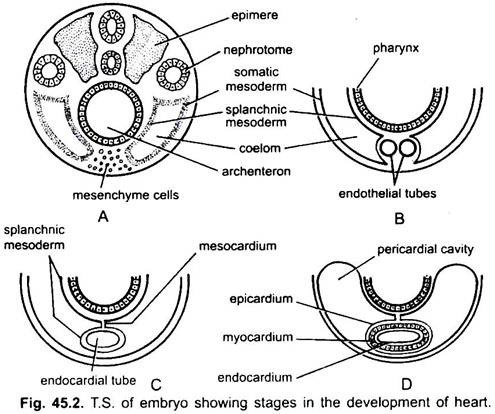
srdce je nepárový orgán, ale jeho původ je bilaterální. V embryu tvoří mezenchym skupinu endokardiálních buněk pod hltanem. Tyto buňky jsou uspořádány tak, aby vytvořily pár tenkých endotelových trubic. Obě endotelové trubice se brzy spojí a vytvoří jednu endokardiální trubici ležící podélně pod hltanem.
splanchnický mezoderm ležící pod endodermem se přehne podélně kolem endokardiální trubice. Tato dvouvrstvá trubice vytvoří srdce, ve kterém splanchnický mezoderm zhoustne a vytvoří myokard nebo svalovou stěnu srdce a vnější tenký epikard nebo viscerální perikard. Endokardiální trubice se stává výstelkou srdce známou jako endokard.
záhyby splanchnického mezodermu se setkávají výše a tvoří hřbetní mezokard, který pozastavuje srdce v coelomu. Brzy se za srdcem vytvoří příčná přepážka, která rozděluje coelom na dvě komory, přední perikardiální dutinu obklopující srdce a zadní břišní dutinu.
srdce je přímá trubice, ale zvětšuje se na délku a stává se tvarem S, protože jeho konce jsou pevné. Vzhled ventilů, zúžení, přepážky v srdci a diferenciální zesílení jeho stěn tvoří tři nebo čtyři komory v srdci.
1. Jednokomorové srdce:
v Amphioxus (primitivní chordát) se pravé srdce nenachází. Část ventrální aorty pod hltanem je svalová a kontraktilní a působí jako srdce.
2. Dvoukomorové Srdce:
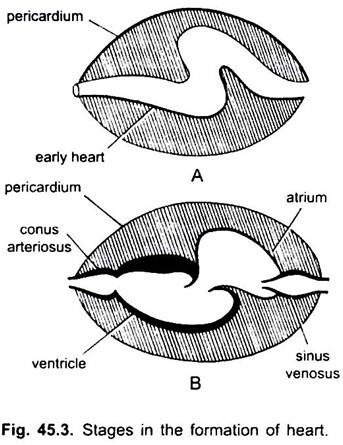
v cyklostomech jsou čtyři komory uspořádané v lineárním pořadí – tenkostěnný sinus venosus, mírně svalová síň (ušnice), svalová komora a svalový konus arteriosus nebo bulbus cordis. Leží v tělní dutině, ve které jsou přítomny i jiné viscerální orgány.
ze čtyř komor odpovídá pouze atrium a komora čtyřem komorám (spárované síně a spárované Komory) vyšších obratlovců. Ve vývoji srdce došlo k mnoha změnám.
Elasmobranchs:
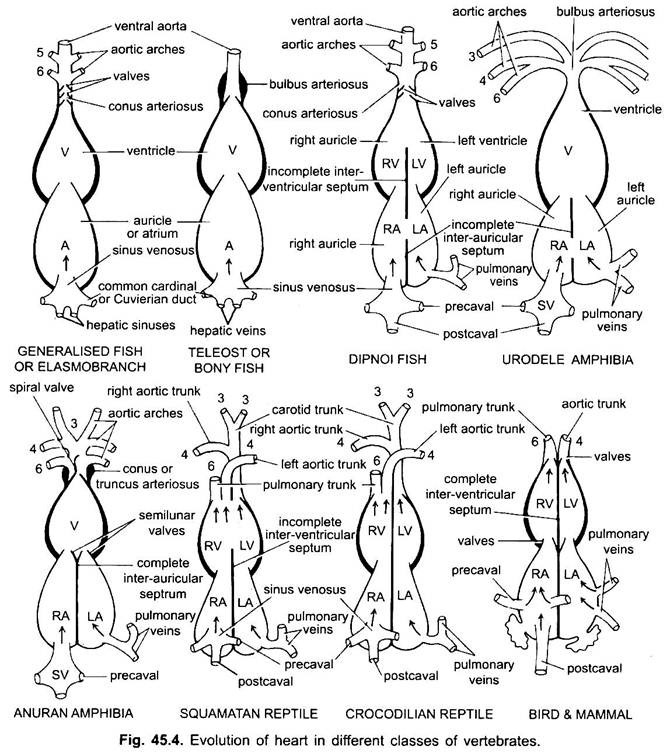
kromě Dipnoi, oběhového systému v rybách od cyklostomů po teleosty, pouze neoxygenovaná krev jde do srdce, odtud je čerpána do žábrů, provzdušňována a poté distribuována do těla. Srdce chrupavčitých dogfish je svalnatá a dorsoventrálně ohnutá trubice ve tvaru písmene S se čtyřmi oddíly v lineární řadě.
jsou to sinus venosus a atrium pro příjem žilní krve a komora a conus arteriosus pro čerpání této krve. Srdce je větvené žilní srdce. Sinus venosus a conus arteriosus jsou doplňkové Komory. Atrium a komora jsou pravé komory, jedná se tedy o 2komorové srdce.
sinus venosus se otevírá dopředu do atria sinuatriální clonou chráněnou dvojicí ventilů. Atrium leží hřbetní k komoře a otevírá se ventrálně do komory prostřednictvím atrioventrikulárního otvoru chráněného dvojicí ventilů. Silnostěnná svalová komora se otevírá do úzkého konus arteriosus obsahujícího ventily ve dvou sériích.
srdce je uzavřeno v perikardiální dutině oddělené od tělesné dutiny příčnou přepážkou. Konus propichuje perikard a stává se spojitým s ventrální aortou. Perikardiální dutina komunikuje s tělní dutinou dvěma perforacemi v příčném septu.
Teleosts:
jejich srdce se podobá srdci clasmobranchů. V teleostech je konus redukován a má jediný pár ventilů. Proximální část ventrální aorty v blízkosti konusu se výrazně zvětšuje a silnostěnná, nazývaná bulbus arteriosus. Je elastický a rozšiřuje se v době komorové kontrakce. Srdce je tedy 2-komorové s jediným oběhem krve.
3. Tříkomorové srdce:
Dipnoi:
v diphoans přepážka rozděluje atrium na pravou a levou komoru. To souvisí s použitím plaveckého močového měchýře jako orgánu dýchání a představuje první krok k rozvoji oběhového systému dvojitého typu, kdy jak okysličená, tak neoxygenovaná krev vstupují do srdce a jsou udržovány odděleně.
krev z pravé ušní trubice plic prochází do pravé komory a je pak čerpána do primitivního plicního plynového močového měchýře plicními tepnami, které se oddělují od šestého páru aortálních oblouků. Okysličená krev se vrací do levé síně prostřednictvím plicních žil, jako jsou obojživelníci.
obojživelníci:
u obojživelníků se hřbetní síň posune před komoru. Sinus venosus se otevírá do pravé síně dorzálně a ne posteriorně. Atrium je zcela rozděleno na pravou a levou komoru a nemá foramen ovale v inter-aurikulární přepážce, která zůstává otevřená v dipnoanech.
hluboké kapsy se vyvíjejí v komorové dutině. Conus arteriosus se dělí na systémové a plicní cévy spirálovým ventilem. U plicních mloků je inter-atriální septa neúplná a plicní žíly chybí.
Reptilia:
u plazů je srdce dále pokročilé. Atrium je vždy zcela odděleno do pravé a levé komory a v mnoha formách je sinus venosus začleněn do stěny pravé síně. Komora je také částečně rozdělena přepážkou u většiny plazů a u aligátorů a krokodýlů je zcela dvoukomorová.
to znamená, že okysličená krev přicházející z plic na levou stranu srdce je v podstatě oddělena od neokysličené krve z těla na pravou stranu. U krokodýlů jsou tedy dva typy krve zcela odděleny a téměř kompletní u jiných plazů, ale k míšení dochází v jiných částech oběhového systému.
embryonální conus arteriosus se rozdělí na tři místo dvou cév:
(i) plicní oblouk nesoucí krev do plic z pravé strany Komory.
(ii) pravá systémová aorta nesoucí krev z levé strany komory do těla prostřednictvím pravého čtvrtého aortálního oblouku.
(iii) levý systém pochází z pravé komory do levého čtvrtého aortálního oblouku.
v místě kontaktu se systémovou aortou z levé komory, dokonce i u krokodýlů, je přítomen otvor mezi těmito dvěma, nazývaný foramen Panizzae, kde může dojít k míšení obou typů krve. Plazové srdce tedy představuje přechodné srdce proti obojživelnému srdci – 2 kompletní ušnice a 2 neúplné komory s malým smícháním krve v pravém a levém systému.
4. Čtyřkomorové srdce:
Aves a Mammalia:
u ptáků je komora zcela rozdělena na dvě, takže srdce je čtyřkomorové (2 ušnice a 2 komory). Existuje úplné oddělení žilní a arteriální krve. Systémová aorta opouští levou komoru a nese krev do hlavy a těla. Zatímco plicní tepna opouští pravou komoru a přenáší krev do plic pro okysličení.
Existuje tedy dvojitý oběh, ve kterém nedochází k žádnému míchání krve. Sinus venosus je zcela začleněn do pravého ušního boltce, který přijímá dva prekavaly a postcaval. Levá ušnice přijímá okysličenou krev plicními žilkami, conus arteriosus chybí, plicní aorta vychází z pravé komory a jediná systémová aorta vychází z levé komory a obě mají ventily na svých základnách.
modifikace aortálních oblouků u obratlovců:
embryonální tepny:
když se srdce tvoří v embryu obratlovců, objeví se krevní céva zvaná ventrální aorta uprostřed ventrálně pod hltanem, která se brzy připojí k conus arteriosus. Ventrální aorta vznikající ze srdce běží dopředu pod hltanem a dělí se dopředu na pár vnějších krčních tepen do hlavy.
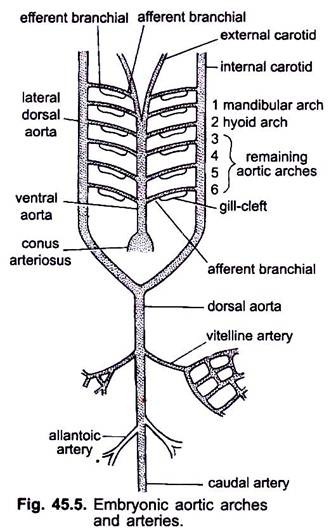
ventrální aorta vydává 6 párů laterálních aortálních oblouků při rovnoběžnosti, které procházejí viscerálními oblouky. Každý aortální oblouk má ventrální aferentní branchiální tepnu nesoucí žilní krev do žábru a dorzální eferentní branchiální tepnu odebírající okysličenou krev z žábru. Eferentní branchiální tepny na obou stranách se spojují dorzálně s laterální dorzální aortou nebo radixovou aortou, která vstupuje do hlavy jako vnitřní krční tepna.
první aortální oblouk je mandibulární aortální oblouk, druhý je hyoidní aortální oblouk, zbývající se nazývají třetí, čtvrtý, pátý a šestý aortální oblouk. Laterální dosální aorta se spojí za hltanem a vytvoří dorzální aortu, která pokračuje uprostřed dorzálně do ocasu jako kaudální tepna.
z dorzální aorty vznikají spárované a nepárové tepny, které zásobují různé orgány těla. V embryu se žloutkovým vakem vzniká pár vitellinových tepen z dorzální aorty a dodává žloutkový vak. U embryí amniot vzniká pár pupečních nebo alantoických tepen z dorzální aorty dodávající krev alantoisům.
u dospělých se vitellinové tepny spojí a vytvoří hlavní mezenterickou tepnu, hlavní část alantoických tepen se ztratí, ale jejich zbytky tvoří hypogastrické nebo vnitřní iliakální tepny.
aortální oblouky u obratlovců:
u různých dospělých obratlovců se arteriální systém jeví jako odlišný, ale jsou postaveny na stejném základním základním plánu. Rozdíl je způsoben rostoucí složitostí srdce v důsledku změny dýchání z žábrů na plíce. V řadě obratlovců dochází k postupnému snižování počtu oblouků aorty.
Cyclostomata:
v Petromyzonu je 7 párů aortálních oblouků. V jiných cyklostomech se liší od 6 párů v Myxinu a 15 párů v Eptatretusu.
ryby:
ačkoli šest je považováno za základní počet aortálních oblouků pro ryby. Toto číslo je sníženo na pět i u žraloků a paprsků se ztrátou prvního páru, mandibulárního aortálního oblouku nebo je reprezentováno eferentní pseudobranchiální tepnou. U většiny kostnatých ryb mizí nebo jsou výrazně sníženy jak mandibulární (i), tak hyoidní (ii) aortální oblouky.
u Polypterus a Dipnoi (plicní ryby) nejsou žábry dobře vyvinuty. Plicní tepna tedy vychází z eferentní části 6. oblouku na obou stranách, která dodává krev do vzduchového močového měchýře nebo plic. V elasmobranchech a Dipnoi má každý oblouk jednu aferentní a dvě eferentní větvové tepny (vytvořené štěpením) v každém žábru. U kostnatých ryb má každá žábra jednu aferentní a jednu eferentní tepnu.
v tetrapodových aortálních obloucích se nerozpadají na aferentní a eferentní části, protože chybí pravé vnitřní žábry. Ve všech tetrapodách zmizí první a druhý oblouk.
Amphibia:
zde aortální oblouky vykazují modifikaci v důsledku ztráty žábrů a vzhledu plic. V urodelech jsou kromě plic přítomny vnější žábry jako dýchací orgány. III, IV, V A VI aortální oblouky jsou přítomny, i když pátý pár je mnohem redukován v siréně, Amphiuma a Necturus. Aortální oblouky nejsou ve vnějších žábrách rozděleny na aferentní a eferentní části, protože větve vznikající z IV, V a VI aortálních oblouků tvoří kapiláry ve vnějších žábrách.
laterální dorzální aorty mezi III a IV aortálními oblouky přetrvávají jako vaskulární spojení, ductus caroticus. VI aortální oblouk tvoří pulmo-kožní oblouk nebo tepnu na obou stranách, která odebírá krev do plic a kůže. Zachovává také spojení s laterální dorzální aortou známou jako ductus arteriosus(kanál Botalli).
v larvě anuranu (žabí pulec) je uspořádání aortálních oblouků podobné dospělému urodele kvůli přítomnosti žábrů. Při metamorfóze, se ztrátou žábrů, I, II a v aortální oblouky úplně zmizí, jsou přítomny pouze III., IV. a Vlth aortální oblouky. Laterální dorzální aorta mezi třetím a čtvrtým aortálním obloukem (ductus caroticus) také zmizí. Třetí aortální oblouk spolu s částí ventrální aorty se tak stává karotidovým obloukem nesoucím okysličenou krev do oblasti hlavy.
čtvrtý aortální oblouk podél jeho laterální dorzální aorty tvoří systémový oblouk. Šestý aortální oblouk se stává pulmokutánním obloukem dodávajícím žilní krev do plic a kůže. Ductus arteriosus zmizí během metamorfózy. Dospělí anurani tedy mají pouze III, IV a VI aortální oblouky. Ty jsou také zachovány amnioty.
Reptilia:
u plazů jsou žábry plně nahrazeny plícemi. Jsou přítomny pouze III, IV a VI aortální oblouky. Při částečném oddělení komory na dvě části se distální část konus arteriosus a celá ventrální aorta rozdělí na tři cévy, tj. Pravý systémový oblouk (IV) vychází z levé komory nesoucí okysličenou krev do karotidového oblouku (III). Levá systémová (IV) a plicní aortae (VI) pocházejí z pravé komory. Levá systémová nese deoxygenovanou nebo smíšenou krev do těla přes dorzální aortu.
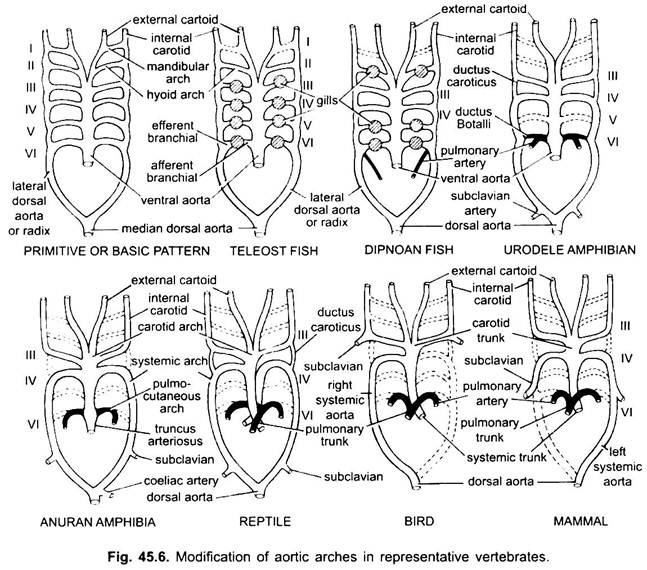
zatímco plicní tepna odvádí deoxygenovanou krev do plic. Ductus caroticus zmizí, ale přetrvává u hadů a některých ještěrek (Uromastix). Ductus arteriosus zmizí u většiny plazů, i když přetrvává ve snížené formě u Sphenodonu a některých želv. Kvůli míchání krve, plazi jsou chladnokrevná zvířata, jako jsou ryby a obojživelníci.
Aves:
u ptáků jsou přítomny oblouky III, IV a VI. Sledují obecný vzorec plazů s určitými rozdíly. Při úplném rozdělení komory na dvě části se konus arteriosus a ventrální aorta rozdělily a vytvořily dvě cévy, systémovou aortu vznikající z levé komory a plicní aortu z pravé komory.
třetí aortální oblouk se zbytky laterální a ventrální aorty tvoří karotidy, které vznikají ze systémové aorty. Čtvrtý aortální oblouk tvoří systémovou aortu pouze na pravé straně. Spojuje se s laterální aortou své vlastní strany a tvoří hřbetní aortu. Část čtvrtého aortálního oblouku levé strany tvoří levou podklíčkovou tepnu, zbytek spolu s laterální dorzální aortou zmizí. Šestý aortální oblouk tvoří plicní aortu. Ductus caroticus a ductus arteriosus zmizí.
Mammalia:
u savců přetrvávají také III, IV a VI aorické oblouky. Komora je zcela rozdělena na dvě části.
konus arteriosus a ventrální aorta se rozdělí na dvě cévy:
(i) systémová aorta vznikající z levé komory a
(ii) plicní aorta z pravé komory. Třetí aortální oblouk se zbytky laterální a ventrální aorty tvoří krční oblouk.
čtvrtý aortální oblouk tvoří systémovou aortu pouze na levé straně, zatímco na pravé straně jeho proximální část tvoří innominovanou a pravou subklavní tepnu, zbytek spolu s laterální dorzální aortou zmizí. Šestý aortální oblouk tvoří plicní aortu. Ductus arteriosus degeneruje, ale v některých přetrvává až do vylíhnutí nebo narození ve snížené formě na levé straně jako tenký ligamentum arteriosum.
Žilní Systém:
Embryonální Žíly:
u všech embryí obratlovců je žilní systém jednoduchý a podobný. Žíly jsou většinou spárovány a symetricky uspořádány. U embryí bez žloutkového vaku se v splanchnickém mezodermu pod střevem vytvoří sub-střevní žíla.
smyčky kolem řitního otvoru a pokračuje posteriorně jako:
(i) kaudální žíla do ocasu. Ve všech embryích, které mají žloutkový vak (ať už obsahuje žloutek nebo ne), vzniká ze žloutkového vaku pár vitellinových žil a spojuje se se zadní částí vyvíjejícího se srdce, které se stává sinusovým venosusem. Ve skutečnosti je fúze těchto vitelline žil zodpovědná za tvorbu srdce u kostnatých ryb, plazů a ptáků. Každá vitellinová žíla na svém zadním konci se připojuje k subintestinální žíle vytvořené stejným způsobem jako u embryí bez žloutkového vaku.
(ii) mezi ledvinami vzniká pár subkardinálních žil a spojuje kaudální žílu.
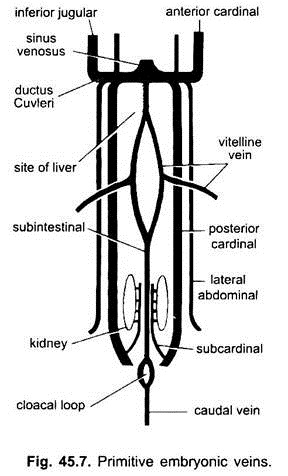
(iii) jsou vytvořeny spárované přední a zadní kardinální žíly, které nesou krev z hlavy a zadní části těla. Přední a zadní kardinální žíly na každé straně se spojují a vytvářejí ductus Cuvieri nebo společnou kardinální žílu, která prochází dovnitř příčnou přepážkou a vstupuje do sinus venosus.
u ryb a mloků (urodeles) pochází dolní jugulární žíla z ventrální strany hlavy, aby se připojila ke společné kardinální žíle. U jiných obratlovců nemá homolog. V amniotes pár laterálních nebo ventrálních břišních žil pochází ze stěny těla, aby vstoupil do společných kardinálních žil.
ryby:
společná kardinální žíla (kanál Cuvier) vstupuje do sinus venosus z každé strany a je tvořena fúzí předních a zadních kardinálů. Krev z hlavy je shromažďována předními kardinály a krev z ledvin a pohlavních žláz je shromažďována zadními kardinály.
také vstupují do kanálků Cuvieru spárované boční břišní žíly, které dostávají krev z stěny těla a spárované přídavky. Renální portální systém se skládá z kaudální žíly a dvou renálních portálních žil umístěných laterálně k ledvinám, které se kapillarizovaly v ledvinách.
jaterní portální systém přenáší krev ze žaludku a střeva a vrací ji do jater, kde po průchodu sérií sinusoidů vstupuje do sinus venosus prostřednictvím párových hapatických žil.
v teleostech chybí laterální břišní žíly, takže krev z subklaviánů, odvodňující prsní přídavky, vstupuje přímo do sinus venosus a krev z iliakálních žil, odvodňující pánevní přídavky, přechází do postkardinálů.
u dipnoanů je přítomna jediná ventrální břišní žíla, pravděpodobně odvozená z fúze laterálních břišních svalů. Tato žíla přijímá krev z ilacs prostřednictvím spárovaných pánevních žil a vstupuje do pravého kanálu Cuvier. Kromě toho, z pravého postkardinálního systému, nové žíly, postclaval dělá jeho vzhled, který má velký význam u vyšších obratlovců. Je spojen s kaudální žílou a prochází vpřed játry do sinusu.
u embryí amniot jsou laterální břišní žíly známé jako pupeční nebo alantoické žíly, protože vypouštějí alantois. Vitelline a pupeční žíly jsou při narození ztraceny, pouze jejich zbytky přetrvávají.
modifikace žil:
stejně jako v případě tepen jsou žíly různých obratlovců uspořádány na stejném základním plánu. Jakékoli změny, které ukazují, sledují sekvenci v řadě obratlovců. Ve svém vývoji prochází žilní systém vyšších obratlovců stádii pozorovanými u embryí nižších forem.
1. Když se tvoří játra, proximální část vitelline žil nebo subintestinálních žil tvoří jaterní žíly mezi játry a srdcem, distální část levé vitelline žíly nebo subintestinální žíly se stává jaterní portální žíla, která tvoří sinusoidy v játrech, což vede ke vzniku jaterního portálního systému přítomného ve všech.
2. Přední kardinální žíly přetrvávají jako vnitřní jugulární žíly.
3. S výjimkou ryb se běžné kardinální žíly stávají prekaválními žilkami, které vstupují do sinus venosus (obojživelníci ,plazi) nebo do pravého ušního boltce(ptáci, savci). V každé přední končetině se vytvoří subklavní žíla, která se připojí k prekavální žíle.
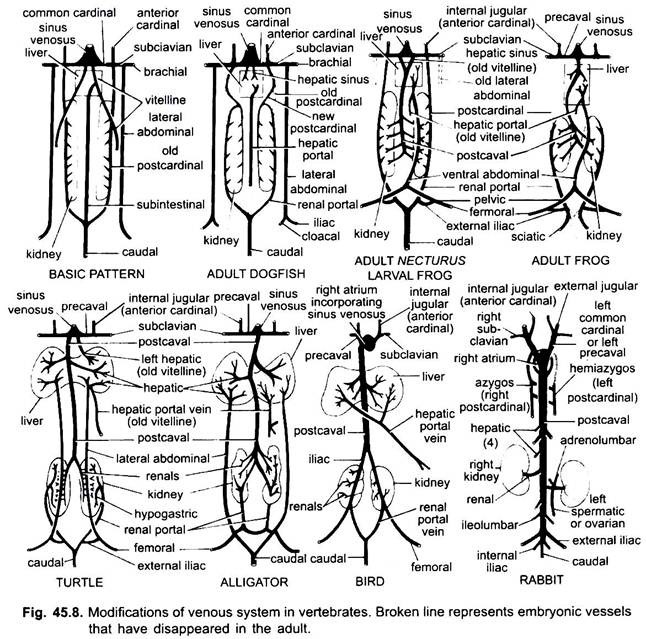
4. Kaudální žíla ztrácí spojení se subintestinálními a subkardinálními žilkami, její přední část se rozdělí na dvě větve, které spojují zadní kardinální žíly u všech kromě savců.
5. Zadní kardinální žíly přetrvávají jako takové u ryb, ale v jiných se každý rozpadne na dvě části, přední část, která zmizí u obojživelníků, plazů a ptáků, ale u savců pravá přední část tvoří azygosovu žílu, levá může tvořit hemiazygos žílu, která ztrácí spojení s prekavální a spojuje azygos žílu příčnou anastomózou; zadní části zadních kardinálních žil, které se připojily k kaudální žíle, se stávají renálními portálními žilami.
renální portální žíla na každé straně tvoří kapiláry v ledvinách a tvoří renální portální systém, který je kompletní u ryb, obojživelníků a plazů, ale u ptáků se výrazně zmenšuje a u savců chybí, protože zadní části zadních kardinálních žil u savců zmizí.
6. Renální portálové žíly ryb sbírají krev pouze z ocasu, ale u obojživelníků, plazů a ptáků také uzurpují žíly z nohou.
7. U některých ryb a všech tetrapod se vitellinové žíly připojují k subkardinální žíle a vytvářejí postkavální žílu. U obojživelníků a plazů se postcavální žíla rozšiřuje na zadní konce ledvin, u ptáků se připojuje k renálním portálním žilám, čímž se snižuje renální portální systém, u savců se postcavální žíla spojuje s žilami z nohou a ocasu, takže systém renálního portálu je zcela eliminován.
8. Dvě boční břišní žíly přetrvávají jako takové u ryb, ale od Dipnoi nahoru se spojí a vytvoří přední břišní žílu, která spojuje jaterní portální žílu poblíž jater, přední břišní žílu, tedy spojuje renální portál a jaterní portální systémy. Iliakální žíla je vytvořena v každé zadní končetině, která spojuje přední břišní žílu. Přední břišní žíla začíná ztrácet svůj význam u plazů, u ptáků je modifikována tak, aby tvořila epigastrické a kokcygeo-mezenterické žíly, u savců zmizí, s výjimkou Tachyglosu.
9. Ve formách dýchajících vzduch vyrůstají plicní žíly z levého ucha a vstupují do plic.
10. Umbilické (alantoické) žíly embrya zmizí, když se plíce stanou funkčními jako respirační orgány.
Tetrapoda:
žilní systém obojživelníků se velmi podobá systému plic, kromě toho, že břišní žíla vstupuje spíše do jaterního portálního systému než do sinus venosus. Přední kardinálové přetrvávají jako vnitřní jugulární žíly u všech dospělých tetrapodů. Dolní jugulární žíly chybí. Společnými kardinály se stávají přední vena cavae nebo precavals, které se připojují k sinus venosus u obojživelníků a plazů.
u ptáků a savců prekavaly přímo vstupují do pravého ucha srdce, protože sinus venosus chybí. U člověka, kočky atd., levý prekaval je ztracen, takže krev levé strany vstupuje do pravého prekavalu brachio-cefalickou větví. U mloků bez plic chybí plicní žíly, protože inter-atriální septa je neúplná.
post kardinální žíly se připojují k kaudální žíle posteriorně a prekavaly dopředu, aby vytvořily společné kardinály v larvální žábě a Nekturu.
u plazů dochází k většímu rozvoji plicních žil a postcavální žíly a ke snížení významu renálního portálního systému, který přivádí krev do ledvin ze zadní části těla.
u ptáků existují dvě funkční prekavální žíly tvořené spojením krční a podklíčkové na každé straně a kompletní post caval. Přijímá krev z končetin prostřednictvím renálních portálů, které procházejí ledvinami, ale nerozpadají se do kapilár a nejsou srovnatelné s renálními portály dolních obratlovců.
u savců může být jedna nebo dvě prekavální žíly. Existuje jediná postcavální žíla, jejíž embryologický vývoj je velmi komplikovaný. Všechny kavální žíly vstupují přímo do pravého ucha, protože sinus venosus je absorbován do stěny této komory srdce v embryonálním životě. Neexistuje žádný renální portálový systém, i když jaterní portálový systém se velmi podobá systému jiných obratlovců.