význam
roste zájem o cirkrna kvůli jejich vlivu na mnoho biologických procesů a nemocí kromě jejich biomarkerového potenciálu. Jsou detekovány hlavně přítomností čtení mapujících jejich backsplicing křižovatku. Nicméně, cirkrna již nejsou jedinými transkripty obsahujícími takovou křižovatku, protože nedávné studie odhalily, že kruhové dna jsou běžné a mohou být přepisovány, což má za následek transkripty, které by napodobovaly cirkrna signál. Proto tento nový typ chimérického přepisu může změnit způsob, jakým se provádí analýza circRNA, a ovlivnit některé z již hlášených výsledků.
jsou kruhové RNA jedinými Chimérickými Přepisy?
kruhová RNA (cirkrna) byla před několika lety znovu objevena jako nekanonicky splétané RNA formy přítomné v různých organismech včetně lidí (Salzman et al., 2012; Jeck et al., 2013). Jedná se o kovalentně uzavřené transkripty vytvořené prostřednictvím události zpětného sestřihu RNA, kde se spojovací dárce následného exonu připojí k akceptoru upstreamového spoje, což vede k kovalentně uzavřeným transkriptům, které jsou charakterizovány přítomností zpětného sestřihu, díky němuž jsou cirkrna odlišitelné od jejich lineárních protějšků (obrázek 1A) (Zhang et al ., 2016; Wilusz, 2018).
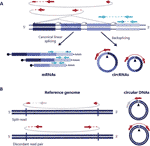
Obrázek 1 detekce obě kruhové RNA a DNAs je založena na čtení překlenujících křižovatku. (A) Backspliced spojení mezi exony a kanonické exon spojování jsou zobrazeny s non-spojité a spojité čáry, respektive vede k cirkrna a mRNA. Párová-end čte klenout backspliced křižovatky jsou zobrazeny červeně a párová-end čte v souladu s lineárním přepisem jsou zobrazeny modře. (B) podobně jako cirkrna jsou kruhové DNAs (eccdna) detekovány na základě strukturně čtených variant v souladu s kruhově znázorněnou červeně .
od jejich znovuobjevení vědecká komunita upozornila na circrna a zkoumala jejich zapojení do několika buněčných procesů ve zdraví a nemoci (Haque a Harries, 2017), jejich potenciální roli biomarkerů (Abu a Jamal, 2016)a jejich regulační funkce (Floris et al ., 2016). Cirkrna jsou nyní známy jako hojné a stabilní v cytosolu a jádru (Salzman et al ., 2012; Jeck et al., 2013; Li et al., 2015) a byly také nalezeny volné v biofluidech (Bahn et al., 2015; Memczak a kol., 2015; Chen a kol., 2018) a v extracelulárních váčcích (Kyoung Mi et al., 2017). Potenciál biomarkerů cirkrna byl intenzivně studován, ve skutečnosti bylo publikováno mnoho případových kontrolních studií hledajících diferencovaně exprimované cirkrna, které by mohly být biomarkery různých onemocnění. K dnešnímu dni byly circrna zapojeny do několika nemocí, včetně rakoviny (Kristensen et al ., 2017; Arnaiz et al., 2018), neurologické poruchy (Akhter, 2018), kardiovaskulární onemocnění (Aufiero a kol., 2019) a nemoci související s imunitou (Iparraguirre et al., 2017; Liu a kol., 2019). Ve stejnou dobu, plně porozumět jejich biogenezi, vlastnosti, funkce, a důsledky v biologii člověka zůstávají otevřenými otázkami pro výzkumné pracovníky v oboru.
ačkoli funkce většiny cirkrna zůstává neznámá, bylo prokázáno, že některé cirkrna mohou působit jako mikroRNA houby, regulující hladiny mikroRNA a jejich aktivitu (Hansen et al ., 2013; Memczak a kol., 2013; Zheng et al., 2016). Podílejí se na regulaci genové exprese regulací transkripce svých rodičovských genů, soutěží s lineárním sestřihem nebo spongingovými proteiny (Ashwal-Fluss et al ., 2014; Li a kol., 2018). Zajímavé je, že studie profilování ribozomů nedávno ukázaly, že cirkrna mohou být také přeloženy Jak in vitro, tak in vivo (Legnini et al ., 2017; Yang a kol., 2017).
hlavním rysem cirkrna a zodpovědným za většinu jejich zvláštních vlastností je jejich kruhovitost. Proto, kromě detekce jejich charakteristické zpětné spojení, testování kruhovitosti těchto molekul, je jedním z klíčových bodů v každé studii circRNA. Nicméně, mnoho studií založilo svůj objev cirkrna na celkové RNA a mohlo tak interpretovat některé lineární chimérické transkripty jako cirkrna, což má za následek falešně pozitivní detekce cirkrna. K obcházení tohoto problému většina studií potvrdila kruhovitost transkriptů nalezených celkovou RNA-seq pomocí rnázy R, Northern blot nebo elektroforetických metod (Jeck a Sharpless, 2014). Tyto ověření kruhovitosti však také někdy odhalily transkripty, které se zdají být lineární, spíše než kruhové, což potvrzuje, že detekce cirkrna počínaje celkovou RNA může vést k některým falešně pozitivním výsledkům. Tyto falešně pozitivní výsledky byly přičítány technickým artefaktům nebo přepisům odvozeným z neobvyklých událostí, jako jsou exonové duplikace nebo transplantační události (Jeck a Sharpless, 2014; Szabo a Salzman, 2016). To znamená, že možnost nalezení pravdivých, biologicky aktivních a funkčních lineárních transkriptů, které obsahují sekvenci ekvivalentní zpětné vazbě (od nynějška nazývané chimérické lineární transkripty), byla poněkud přehlížena, protože zdroj takové lineární RNA nebyl znám pro zdravé buňky.
kruhové dna jako zdroj chimérických lineárních transkriptů
většina lidského genomu je organizována v lineárních chromozomech, nicméně některé výjimky jsou již dlouho přijímány, jako je mitochondriální DNA, a chromozomální aberace, jako jsou DNA kruhy nesoucí onkogeny (např., 1991; Nathanson et al., 2014; Turner a kol., 2017) a kruhové chromozomy (Tümer et al., 2004). Až donedávna byly různé kruhové dna, jako jsou mikrodny (Shibata et al ., 2012) nebo extrachromozomální kruhové DNAs (eccdna)byly také nalezeny z velkých částí různých eukaryotických genomů včetně člověka a kvasinek (Møller et al ., 2015; Kumar a kol., 2017; Møller a kol., 2018).
kruhové dna jsou vytvořeny, když jsou dva konce lineární DNA spojeny dohromady, což vede ke spojení podobnému backspliced křižovatce na circrna běžně nazývaném breakpoint junction, který je detekován na základě strukturně čtených variant v souladu s cirkulační událostí (Gresham et al., 2010; Møller et al., 2018; Prada-Luengo a kol., 2019) (obrázek 1B). Obvykle se pohybují od stovky bází po megabázové kruhy a mohou obsahovat plné exony a geny (Shibata et al ., 2012; Møller a kol., 2015; Kumar a kol., 2017; Turner a kol., 2017; Møller a kol., 2018) a zatímco některé oblasti genomu se častěji vyskytují na kruhové DNA (Sinclair a Guarente, 1997;; Møller et al., 2016; Turner a kol., 2017; Møller a kol., 2018) se zdá, že většina kruhové DNA se vyskytuje náhodně (Shibata et al ., 2012; Møller a kol., 2015; Kumar a kol., 2017; Møller a kol., 2018).
zajímavé je, že v nedávném článku Møller et al. identifikovány tisíce eccdna v leukocytech a svalových buňkách u zdravých kontrol. S myšlenkou prozkoumat, zda lze eccdna přepsat, byla také sekvenována knihovna mRNA ze svalové tkáně a analyzována na transkripční události napříč zlomovým bodem detekované eccDNA, která našla několik shod (Møller et al ., 2018). Toto zjištění naznačuje, že kruhová DNA ve zdravé tkáni je transkribována, což vede k lineárním a polyadenylovaným transkriptům, které budou nést sekvenci ekvivalentní zpětné sekvenci cirkrna (Møller et al., 2018) (Obrázek 2).
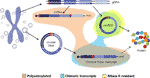
Obrázek 2 grafické znázornění různých transkriptů pocházejících buď z genomové DNA, nebo z kruhové DNA. Exony jsou zbarveny fialově a backspliced křižovatky nebo chimérické křižovatky jsou zobrazeny červeně. Polyadenylované, chimérické spojení obsahující a rnáza R rezistentní transkripty jsou zvýrazněny oranžově, modrá a žlutá.
transkripční důkazy kruhových DNA, spolu s jejich hojností, nás vedou k tomu, že kruhové dna by mohly být přirozeným zdrojem podstatného množství lineárních RNA nesoucích chimérické křižovatky. V mnoha případech, tyto chimérické křižovatky mohou být nerozeznatelné od backsplicing křižovatek circrna, a proto by mohly být matoucí faktory ve studiích circRNA. V následujících odstavcích vysvětlíme údaje podporující tento návrh.
detekce circRNA: Vše, co se třpytí, není zlato
jak již bylo dříve zavedeno, circrna jsou tvořeny i když nekanonická sestřihová událost zvaná backsplicing. Přepisy vyplývající z této události backsplicing mají kovalentně uzavřenou strukturu smyčky ani s polaritou 5 ‚-3‘, ani s polyadenylovaným ocasem a co je důležitější, jsou charakterizovány přítomností kódovaného exonového řádu vzhledem k lineárnímu přepisu (Zhang et al., 2016; Wilusz, 2018). Tento kódovaný exon pořadí je patrné v backspliced křižovatce, která spojuje 5 ‚downstream sekvence s upstream 3‘ sekvence. Všechny detekční algoritmy circRNA tedy využívají přítomnost zpětně spojovaných spojů jako diagnostického prvku pro identifikaci circRNA (obrázek 1A).
byly upraveny různé metody pro detekci těchto zpětně spojovaných spojů. Komerční pole obsahující sondy zaměřené na tyto backspliced oblasti byly široce používány ve studiích screeningu biomarkerů (Iparraguirre et al ., 2017; Liu a kol., 2017; Sui a kol., 2017; Li a kol., 2018). Následná validace je často také založena na zesílení zpětných spojů pomocí divergentních primerů (Panda a Gorospe, 2018). Mnoho dalších článků provedlo vysoce výkonnou sekvenční analýzu, která překonává jedno z hlavních omezení polí, které umožňuje detekovat nejen anotované cirkrna, ale také De novo RNA circularizační události z genomických oblastí, kde předchozí studie nebyly anotovány žádnou circRNA. Bylo vyvinuto několik bioinformatických potrubí pro detekci cirkrna v datových sadách RNA-Seq, ale všechny jsou založeny na přítomnosti přečtených přechodů přes spojovací spoje a nalezení nejspolehlivějšího je pro bioinformatiky stále výzvou (Hansen et al ., 2016; Hansen, 2018; Prada-Luengo a kol., 2019).
pro detekci cirkrna v RNA-seq datech lze použít dva hlavní přístupy. Za prvé, mnoho studií circRNA RNA – seq je založeno na vzorcích ošetřených Rnázou R, aby se vyčerpaly všechny lineární RNA před sekvenováním. Přestože je tento přístup speciálně navržen pro detekci cirkrna, stojí za zmínku, že degradace Rnázy R je variabilní, že existují vzácné případy rnázy R rezistentních lineárních RNA a rnázy R citlivých cirkrna (Szabo a Salzman, 2016) a že tato léčba vyžaduje vysoký vstup RNA, který by mohl být pro některé tkáně omezující. Jiné studie circRNA se rozhodly sekvenovat buď celkovou, ribozomálně ochuzenou (ribo -) nebo nepolyadenylovanou (polyA -) RNA, kde lze nalézt lineární i kruhové RNA (Salzman et al., 2012; Memczak a kol., 2013; Broadbent a kol., 2015; Lu et al., 2015; Memczak a kol., 2015). Tento přístup se vyhýbá použití Rnázy R, která snižuje množství RNA potřebné pro sekvenování a umožňuje studovat expresi jiných typů RNA ze stejné datové sady. Bylo prokázáno, že s dobrou hloubkou a kvalitou sekvenování a pečlivou analýzou dat mohou být skutečné cirkrna detekovány z celkového sekvenování RNA (Wang et al ., 2017) je však v tomto druhém přístupu zapotřebí pozdější potvrzení kruhovitosti pro cirkrna.
s objevem lineárních chimérických RNA transkribovaných z kruhových dna již cirkrna nejsou jedinými transkripty s chimérickými spoji. Proto, je nanejvýš důležité si uvědomit, že zatímco první přístup významně obohatí vzorek RNA v cirkrna tak, že většina detekovaných chimérických spojů bude odpovídat skutečným cirkrna, druhý by mohl přeceňovat počet circRNA transkriptů přiřazením circrna signál přicházející z obou cirkrna a lineární chimérické transkripty přepsané z kruhových DNAs. V důsledku toho, s přihlédnutím ke koexistenci cirkrna a lineárních chimérických transkriptů, nabývá na důležitosti potřeba testů cirkulárnosti a testů funkčnosti a je třeba věnovat zvláštní pozornost nejen experimentálním, ale také výpočetním metodám, aby se zabránilo záměně chimérických transkriptů z kruhových DNAs s cirkrna tvořenými backsplicingem.
diskuse
cirkrna pole je stále v rané fázi, nicméně cirkrna se již ukázaly jako úžasné molekuly, které se podílejí na mnoha procesech, s velkým potenciálem biomarkerů a které mohou také změnit způsob, jakým chápeme transkripční a translační procesy. Z těchto důvodů získávají pozornost a oblast circRNA je v současné době jedním z nejaktivnějších oblastí výzkumu RNA. Stále však existuje mnoho konfliktů, kontroverzí a otevřených otázek (Li, 2019), o kterých je třeba diskutovat.
v této zprávě a ve světle nedávných pokroků v oblasti kruhové DNA chceme poukázat na transkripci z extrachromozomální kruhové DNA jako na jeden z hlavních přírodních zdrojů lineárních transkriptů se zpětně spojenými signály, které by mohly interferovat s daty circRNA (Møller et al., 2018). Od této chvíle bychom kromě technických artefaktů, duplikací a transplantačních událostí, které by mohly vést k falešným pozitivům při detekci circRNA, měli vzít v úvahu také existenci tohoto nového typu chimérických přepisů. Proto jsou cirkulační testy a funkční testy důležitější než kdy jindy.
v každém případě by tyto chimérické lineární transkripty neměly být považovány pouze za pouhý matoucí faktor pro studie circRNA. Navzdory technickým důsledkům pro charakterizaci circRNA, existence těchto circRNA podobných chimérických lineárních molekul RNA pocházejících z eccdna přidává nový typ molekuly do stále rostoucího seznamu RNA a rozšiřuje naši vizi o složitosti transkriptomu a jeho regulaci. Kromě toho by tyto lineární molekuly RNA pocházející z eccDNA mohly také představovat funkce podobné circRNA, včetně regulačních funkcí nebo potenciálu, který má být přeložen. Genové produkty z transkriptů eccDNA by mohly potenciálně přispívat k fenotypu somatických buněk a tkání, jak je uvedeno u kvasinek (Gresham et al., 2010; Demeke et al., 2015). V této rodící se oblasti je však zapotřebí více dat a výzkumu, aby se začalo poškrábat povrch ledovce.
Autor příspěvky
LI, IP-L, BR, and do napsal papír.
financování
tato studie byla financována Instituto de Salud Carlos III prostřednictvím projektu „PI17/00189“ (spolufinancováno Evropským fondem pro regionální rozvoj / Evropským sociálním fondem) „investice do vaší budoucnosti“). IP-L A BR byly podporovány dánskou Radou pro nezávislý výzkum, 6108-00171B a LI byl podporován Ministerstvem školství baskické vlády .
střet zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.